
 | ||
Pleistocene extinctions
The Quaternary period saw the extinctions of numerous predominantly larger, especially megafaunal, species, many of which occurred during the transition from the Pleistocene to the Holocene epoch. However, this extinction wave did not stop at the end of the Pleistocene, but continued, especially on isolated islands, in human-caused extinctions, although there is debate as to whether these should be considered separate events or part of the same event. Among the main causes hypothesized by paleontologists are natural climate change and overkill by humans, who appeared during the Middle Pleistocene and migrated to many regions of the world during the Late Pleistocene and Holocene. A variant of the latter possibility is the second-order predation hypothesis, which focuses more on the indirect damage caused by overcompetition with nonhuman predators. The spread of disease is also discussed as a possible reason.
Contents
- Pleistocene extinctions
- The Pleistocene or Ice Age extinction event
- Africa and Asia
- The Pacific Australasia and Oceania
- Europe and Northern Eurasia
- North America and the Caribbean
- South America
- Later extinctions
- Hunting hypothesis
- Overkill hypothesis
- Objections to the hunting hypothesis
- Climate change hypothesis
- Increased temperature
- Arguments against the temperature hypothesis
- Increased continentality affects vegetation in time or space
- Vegetation changes geographic
- Rainfall changes time
- Arguments against the continentality hypotheses
- Arguments against both climate change and overkill
- Theory
- Arguments against the hyperdisease hypothesis
- Scenario
- Support
- Second order predation and other theories
- Arguments against the second order predation hypothesis
- Arguments against the second order predation plus climate hypothesis
- Comet hypothesis
- Arguments against the comet hypothesis
- References
The Pleistocene or Ice Age extinction event
The Late Pleistocene extinction event saw the extinction of many mammals weighing more than 40 kg.
The extinctions in the Americas entailed the elimination of all the larger (over 1000 kg) mammalian species of South American origin, including those that had migrated north in the Great American Interchange. Only in North America, South America, and Australia, did the extinction occur at family taxonomic levels or higher.
There are three main hypotheses concerning the Pleistocene extinction:
There are some inconsistencies between the current available data and the prehistoric overkill hypothesis. For instance, there are ambiguities around the timing of sudden extinctions of Australian megafauna. Biologists note that comparable extinctions have not occurred in Africa and South or Southeast Asia, where the fauna evolved with hominids. Post-glacial megafaunal extinctions in Africa have been spaced over a longer interval.
Evidence supporting the prehistoric overkill hypothesis includes the persistence of certain island megafauna for several millennia past the disappearance of their continental cousins. Ground sloths survived on the Antilles long after North and South American ground sloths were extinct. The later disappearance of the island species correlates with the later colonization of these islands by humans. Similarly, dwarf woolly mammoths died out on remote Wrangel Island 1,000 years after their extinction on the mainland. Steller's sea cows also persisted in seas off the isolated and uninhabited Commander Islands for thousands of years after they had vanished from the continental shores of the north Pacific.
Alternative hypotheses to the theory of human responsibility include climate change associated with the last glacial period and the Younger Dryas event, as well as Tollmann's hypothetical bolide, which claim that the extinctions resulted from bolide impact(s). Such a scenario has been proposed as a contributing cause of the 1,300 year cold period known as the Younger Dryas stadial. This impact extinction hypothesis is still in debate due to the exacting field techniques required to extract minuscule particles of extra terrestrial impact markers such as Iridium at a high resolution from very thin strata in a repeatable fashion, as is necessary to conclusively distinguish the event peak from the local background level of the marker. The debate seems to be exacerbated by infighting between the Uniformitarianism camp and the Catastrophism camp.
Africa and Asia
The Old World tropics have been relatively spared by Pleistocene extinctions. Africa and southern Asia are the only regions that have terrestrial mammals weighing over 1000 kg today. However, during the early, middle and late Pleistocene some large animal forms disappeared from these regions without being replaced by comparable successor species. Climate change has been cited as most likely causing the extinctions in Southeast Asia.
Megafauna that disappeared in Africa or Asia during the Pleistocene include:
Megafauna that disappeared in Africa and/or Asia during the Late Pleistocene:
The Pacific (Australasia and Oceania)
In Australia, the sudden spate of extinctions occurred earlier than in the Americas. Most evidence points to the period immediately after the first arrival of humans—thought to be a little under 50,000 years ago—but scientific argument continues as to the exact date range. In the rest of the Pacific (New Guinea, New Caledonia, the rest of Australasia and Oceania) although in some respects far later, endemic fauna also usually perished quickly upon the arrival of humans in the late Pleistocene and early Holocene This section does not include any spate of extinctions post 1000 BCE (e.g. subatlantic New Zealand or Hawaii).
The extinctions in the Pacific included:
Some extinct megafauna, such as the bunyip-like Diprotodon, may remain in folk memory or be the sources of cryptozoological legends.
Europe and Northern Eurasia
During the Late Pleistocene, this region was characterised by a great diversity and dynamism of biomes, including the warm climes of the Mediterranean basin, open temperate woodlands, arid plains, mountainous heathland and steppe-tundra which extended to Spain, all of which were subject to the severe climatic fluctuations of stadials. However, Europe, and in particular northern Eurasia, being defined as typically far colder and drier than today, was largely hegemonized by the mammoth steppe, an ecosystem dominated by palatable high-productivity grasses, herbs and willow shrubs. This supported an extensive biota of grassland fauna, and stretched eastwards from Spain in the Iberian Peninsula to the Yukon in modern day Canada.The area was populated by many species of grazers which assembled in large herds similar in size to those in Africa today. Populous species that roamed the great grasslands included the woolly mammoth, woolly rhino, steppe bison, horse, muskox, reindeer and saiga antelope. Carnivores included Eurasian cave lion, Homotherium, cave hyena, grey wolf, dhole, and Arctic fox.
At the edges of these large stretches of grassland could be found more shrub-like terrain and dry conifer forests (similar to taiga). The browsing collective of megafauna included woolly rhinoceros, moose, wapiti, and camels. Brown bears, wolverines, cave bears, lynx, tigers, leopards, and red foxes also inhabited this biome. The more mountainous terrain was occupied by several species of mountain-going animals like the snow sheep, chamois, mouflon, snow leopards and leopards.
There were a number of glacial and inter-glacial periods in this geological age- during the glacial periods, there were large drops in sea level, so the coast-line was greatly modified. The final major cold spell occurred from 25,000 to 18,000 years ago and is known as the Last Glacial Maximum when the Fenno-Scandinavian ice sheet covered much of northern Europe while the Alpine ice sheet occupied significant parts of central-southern Europe. However, there were several less cold periods after this. The epoch ends with the Younger Dryas when there were small ice sheets in Scotland and Scandinavia.
(80,000–4,000 years ago)
Many species extant today were present in areas either far to the south or west of their contemporary ranges- for example, all the arctic fauna on this list inhabited regions as south as the Iberian Peninsula at various stages of the Late Pleistocene. Recently extinct organisms are noted as †. Species extirpated from significant portions of or all former ranges in Europe and Northern Eurasia during the Quaternary Extinction Event include-
North America and the Caribbean
See also: List of North American animals extinct in the Holocene
During the last 60,000 years, including the end of the last glacial period, approximately 51 genera of large mammals have become extinct in North America. Of these, many genera extinctions can be reliably attributed to a brief interval of 11,500 to 10,000 radiocarbon years before present, shortly following the arrival of the Clovis people in North America. Most other extinctions are poorly constrained in time, though some definitely occurred outside of this narrow interval. In contrast, only about half a dozen small mammals disappeared during this time. Previous North American extinction pulses had occurred at the end of glaciations, but not with such an ecological imbalance between large mammals and small ones. (Moreover, previous extinction pulses were not comparable to the Quaternary extinction event; they involved primarily species replacements within ecological niches, while the latter event resulted in many ecological niches being left unoccupied.) The megafaunal extinctions include forty one genera of herbivores (H), and twenty carnivores (C). North American extinctions included:
The survivors are in some ways as significant as the losses: bison (H), grey wolf (C), lynx (C), grizzly bear (C), American black bear (C), deer (e.g. caribou, moose, waipiti (elk), Odocoileus sp.) (H), pronghorn (H), white-lipped peccary (H), muskox (H), bighorn sheep (H), and mountain goat (H); the list of survivors also include species which were extirpated during the Quaternary extinction event, but recolonised at least part of their ranges during the mid-holocene from South American relict populations, such as the cougar (C), jaguar (C), collared peccary (H), ocelot (C), margay (C), and jaguarundi (C). All save the pronghorns descended from Asian ancestors that had evolved with human predators. Pronghorns are the second fastest land mammal (after the cheetah), which may have helped them elude hunters. More difficult to explain in the context of overkill is the survival of bison, since these animals first appeared in North America less than 240,000 years ago and so were geographically removed from human predators for a sizeable period of time. Because ancient bison evolved into living bison, there was no continent-wide extinction of bison at the end of the Pleistocene (although the genus was regionally extirpated in many areas). The survival of bison into the Holocene and recent times is therefore inconsistent with the overkill scenario. By the end of the Pleistocene, when humans first entered North America, these large animals had been geographically separated from human hunters for more than 200,000 years. Given this enormous span of geologic time, bison would almost certainly have been very nearly as naive as native North American large mammals.
The culture that has been connected with the wave of extinctions in North America is the paleo-Indian culture associated with the Clovis people (q.v.), who were thought to use spear throwers to kill large animals. The chief criticism of the "prehistoric overkill hypothesis" has been that the human population at the time was too small and/or not sufficiently widespread geographically to have been capable of such ecologically significant impacts. This criticism does not mean that climate change scenarios explaining the extinction are automatically to be preferred by default, however, any more than weaknesses in climate change arguments can be taken as supporting overkill. Some form of a combination of both factors could be plausible, and overkill would be a lot easier to achieve large-scale extinction with an already dying population due to climate change.
Lack of tameable megafauna was perhaps one of the reasons why Amerindian civilizations evolved differently from Old World ones. Critics have disputed this by arguing that llamas, alpacas, and bison were domesticated.
South America
South America had been isolated, an island continent, for many millions of years, and it had a wide range of fauna found nowhere else, though many of them became extinct during the Great American Interchange about 3 million years ago, such as the Phorusrhacidae and Saparassodonta families. Those that survived the interchange included the ground sloths, glyptodonts, pampatheres, and notoungulates; all managed to extend their range to North America. In the Pleistocene, South America remained largely unglaciated except for increased mountain glaciation in the Andes, with the megafauna were marginally affected. At the start of the Holocene, all the preeminent genera of megafauna became extinct. Their smaller relatives remain, including anteaters, tree sloths, armadillos; New World marsupials: opossums, shrew opossums, and the monito del monte (actually more related to Australian marsupials). Today the largest land mammals remaining in South America are the wild camels of the Lamini group, such as the guanacos and vicuñas, and the Tapirus genus, of which Baird's tapir can reach up to 400 kg. Other notable surviving large fauna are peccaries, marsh deer (Capreolinae), spectacled bears, maned wolves, pumas, ocelots, jaguars, rheas, emerald tree boas, boa constrictors, anacondas, american crocodiles, caimans, and giant rodents such as capybaras.
Later extinctions
There is no general agreement on where the Holocene, or anthropogenic, extinction begins, and the Quaternary extinction event which includes climate change resulting in the end of the last ice age ends, or if they should be considered separate events at all. Some have suggested that anthropogenic extinctions may have begun as early as when the first modern humans spread out of Africa between 100,000 and 200,000 years ago, which is supported by rapid megafaunal extinction following recent human colonisation in Australia, New Zealand and Madagascar, in a similar way that any large, adaptable predator moving into a new ecosystem would. In many cases, it is suggested even minimal hunting pressure was enough to wipe out large fauna, particularly on geographically isolated islands. Only during the most recent parts of the extinction have plants also suffered large losses.
Overall, the Holocene extinction can be characterised by the human impact on the environment. The Holocene extinction continues into the 21st century, with overfishing, ocean acidification and the amphibian crisis being a few broader examples of an almost universal, cosmopolitan decline of biodiversity.
Hunting hypothesis
The hunting hypothesis suggests that humans hunted megaherbivores to extinction, which in turn caused the extinction of carnivores and scavengers which had preyed upon those animals. Therefore, this hypothesis holds Pleistocene humans responsible for the megafaunal extinction. One variant, known as blitzkrieg, portrays this process as relatively quick. Some of the direct evidence for this includes: fossils of some megafauna found in conjunction with human remains, embedded arrows and tool cut marks found in megafaunal bones, and European cave paintings that depict such hunting. Biogeographical evidence is also suggestive: the areas of the world where humans evolved currently have more of their Pleistocene megafaunal diversity (the elephants and rhinos of Asia and Africa) compared to other areas such as Australia, the Americas, Madagascar and New Zealand without the earliest humans. A picture arises of the megafauna of Asia and Africa evolving alongside humans, learning to be wary of them, and in other parts of the world the wildlife appearing ecologically naive and easier to hunt. This is particularly true of island fauna, which display a disastrous lack of fear of humans. Of course, it is impossible to demonstrate this naïveté directly in ancient fauna.
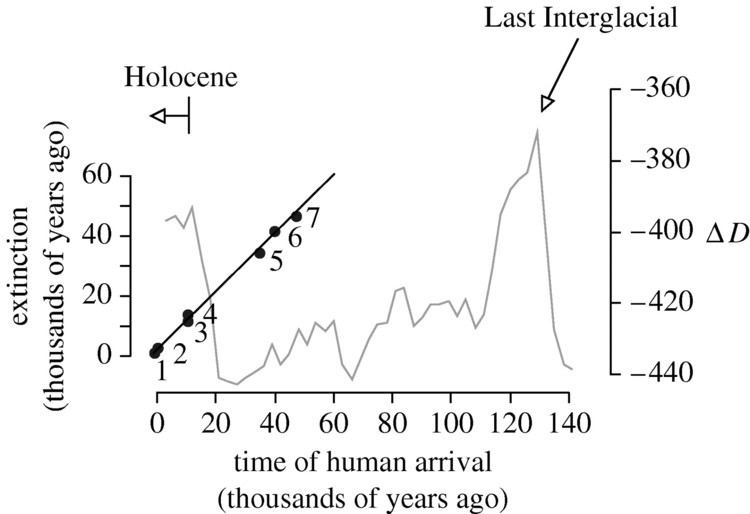
Circumstantially, the close correlation in time between the appearance of humans in an area and extinction there provides weight for this scenario. For example, the woolly mammoth survived on islands despite changing climatic conditions for thousands of years after the end of the last glaciation, but they died out when humans arrived around 1700 BC. The megafaunal extinctions covered a vast period of time and highly variable climatic situations. The earliest extinctions in Australia were complete approximately 50,000 BP, well before the last glacial maximum and before rises in temperature. The most recent extinction in New Zealand was complete no earlier than 500 BP and during a period of cooling. In between these extremes megafaunal extinctions have occurred progressively in such places as North America, South America and Madagascar with no climatic commonality. The only common factor that can be ascertained is the arrival of humans. This phenomenon appears even within regions. The mammal extinction wave in Australia about 50,000 years ago coincides not with known climatic changes, but with the arrival of humans. In addition, large mammal species like the giant kangaroo Protemnodon appear to have succumbed sooner on the Australian mainland than on Tasmania, which was colonised by humans a few thousand years later.
Worldwide, extinctions seem to follow the migration of humans and to be most severe where humans arrived most recently and least severe where humans originated — in Africa (see figure "March of Man" below). This suggests that prey animals and human hunting ability evolved together, so the animals evolved avoidance techniques. As humans migrated throughout the world and became more and more proficient at hunting, they encountered animals that had evolved without the presence of humans. Lacking the fear of humans that African animals had developed, animals outside of Africa were easy prey for human hunting techniques. It also suggests that this is independent of climate change.
Extinction through human hunting has been supported by archaeological finds of mammoths with projectile points embedded in their skeletons, by observations of modern naïve animals allowing hunters to approach easily and by computer models by Mosimann and Martin, and Whittington and Dyke, and most recently by Alroy.
A 2015, a study supported the hypothesis further by running several thousand scenarios that correlated the time windows in which each species is known to have become extinct with the arrival of humans on different continents or islands. This was compared against climate reconstructions for the last 90,000 years. The researchers found correlations of human spread and species extinction indicating that the human impact was the main cause of the extinction, while climate change exacerbated the frequency of extinctions. The study, however, found an apparently low extinction rate in the fossil record of mainland Asia.
Overkill hypothesis
The overkill hypothesis, a variant of the hunting hypothesis, was proposed 40 years ago by Paul S. Martin, Professor of Geosciences Emeritus at the Desert Laboratory of the University of Arizona.
Objections to the hunting hypothesis
The major objections to the theory are as follows:
Climate change hypothesis
At the end of the 19th and beginning of the 20th centuries, when scientists first realized that there had been glacial and interglacial ages, and that they were somehow associated with the prevalence or disappearance of certain animals, they surmised that the termination of the Pleistocene ice age might be an explanation for the extinctions.
Critics object that since there were multiple glacial advances and withdrawals in the evolutionary history of many of the megafauna, it is rather implausible that only after the last glacial would there be such extinctions. However, this criticism is rejected by a recent study indicating that terminal Pleistocene megafaunal community composition may have differed markedly from faunas present during earlier interglacials, particularly with respect to the great abundance and geographic extent of Pleistocene Bison at the end of the epoch. This suggests that the survival of megafaunal populations during earlier interglacials is essentially irrelevant to the terminal Pleistocene extinction event, because bison were not present in similar abundance during any of the earlier interglacials.
Some evidence weighs against climate change as a valid hypothesis as applied to Australia. It has been shown that the prevailing climate at the time of extinction (40,000–50,000 BP) was similar to that of today, and that the extinct animals were strongly adapted to an arid climate. The evidence indicates that all of the extinctions took place in the same short time period, which was the time when humans entered the landscape. The main mechanism for extinction was probably fire (started by humans) in a then much less fire-adapted landscape. Isotopic evidence shows sudden changes in the diet of surviving species, which could correspond to the stress they experienced before extinction.
Evidence in Southeast Asia, in contrast to Europe, Australia, and the Americas, suggests that climate change and an increasing sea level were significant factors in the extinction of several herbivorous species. Alterations in vegetation growth and new access routes for early humans and mammals to previously isolated, localized ecosystems were detrimental to select groups of fauna.
Some evidence obtained from analysis of the tusks of mastodons from the American Great Lakes region appears inconsistent with the climate change hypothesis. Over a span of several thousand years prior to their extinction in the area, the mastodons show a trend of declining age at maturation. This is the opposite of what one would expect if they were experiencing stresses from deteriorating environmental conditions, but is consistent with a reduction in intraspecific competition that would result from a population being reduced by human hunting.
Increased temperature
The most obvious change associated with the termination of an ice age is the increase in temperature. Between 15,000 BP and 10,000 BP, a 6 °C increase in global mean annual temperatures occurred. This was generally thought to be the cause of the extinctions.
According to this hypothesis, a temperature increase sufficient to melt the Wisconsin ice sheet could have placed enough thermal stress on cold-adapted mammals to cause them to die. Their heavy fur, which helps conserve body heat in the glacial cold, might have prevented the dumping of excess heat, causing the mammals to die of heat exhaustion. Large mammals, with their reduced surface area-to-volume ratio, would have fared worse than small mammals.
Arguments against the temperature hypothesis
More recent research has demonstrated that the annual mean temperature of the current interglacial that we have seen for the last 10,000 years is no higher than that of previous interglacials, yet some of the same large mammals survived similar temperature increases. Therefore, warmer temperatures alone may not be a sufficient explanation.
In addition, numerous species such as mammoths on Wrangel Island and St. Paul Island survived in human-free refugia despite changes in climate. This would not be expected if climate change were responsible (unless their maritime climates offered some protection against climate change not afforded to coastal populations on the mainland). Under normal ecological assumptions island populations should be more vulnerable to extinction due to climate change because of small populations and an inability to migrate to more favorable climes.
Increased continentality affects vegetation in time or space
Other scientists have proposed that increasingly extreme weather—hotter summers and colder winters—referred to as "continentality", or related changes in rainfall caused the extinctions. The various hypotheses are outlined below.
Vegetation changes: geographic
It has been shown that vegetation changed from mixed woodland-parkland to separate prairie and woodland. This may have affected the kinds of food available. Shorter growing seasons may have caused the extinction of large herbivores and the dwarfing of many others. In this case, as observed, bison and other large ruminants would have fared better than horses, elephants and other monogastrics, because ruminants are able to extract more nutrition from limited quantities of high-fiber food and better able to deal with anti-herbivory toxins. So, in general, when vegetation becomes more specialized, herbivores with less diet flexibility may be less able to find the mix of vegetation they need to sustain life and reproduce, within a given area.
Rainfall changes: time
Increased continentality resulted in reduced and less predictable rainfall limiting the availability of plants necessary for energy and nutrition. Axelrod and Slaughter have suggested that this change in rainfall restricted the amount of time favorable for reproduction. This could disproportionately harm large animals, since they have longer, more inflexible mating periods, and so may have produced young at unfavorable seasons (i.e., when sufficient food, water, or shelter was unavailable because of shifts in the growing season). In contrast, small mammals, with their shorter life cycles, shorter reproductive cycles, and shorter gestation periods, could have adjusted to the increased unpredictability of the climate, both as individuals and as species which allowed them to synchronize their reproductive efforts with conditions favorable for offspring survival. If so, smaller mammals would have lost fewer offspring and would have been better able to repeat the reproductive effort when circumstances once more favored offspring survival.
Arguments against the continentality hypotheses
Critics have identified a number of problems with the continentality hypotheses.
The extinction of the megafauna could have caused the disappearance of the mammoth steppe. Alaska now has low nutrient soil unable to support bison, mammoths, and horses. R. Dale Guthrie has claimed this as a cause of the extinction of the megafauna there; however, he may be interpreting it backwards. The loss of large herbivores to break up the permafrost allows the cold soils that are unable to support large herbivores today. Today, in the arctic, where trucks have broken the permafrost grasses and diverse flora and fauna can be supported. In addition, Chapin (Chapin 1980) showed that simply adding fertilizer to the soil in Alaska could make grasses grow again like they did in the era of the mammoth steppe. Possibly, the extinction of the megafauna and the corresponding loss of dung is what led to low nutrient levels in modern-day soil and therefore is why the landscape can no longer support megafauna.
Arguments against both climate change and overkill
It may be observed that neither the overkill nor the climate change hypotheses can fully explain events: browsers, mixed feeders and non-ruminant grazer species suffered most, while relatively more ruminant grazers survived. However, a broader variation of the overkill hypothesis may predict this, because changes in vegetation wrought by anthropogenic fire preferentially selects against browse species.
Theory
The Hyperdisease Hypothesis attributes the extinction of large mammals during the late Pleistocene to indirect effects of the newly arrived aboriginal humans. The Hyperdisease Hypothesis proposes that humans or animals traveling with them (e.g., chickens or domestic dogs) introduced one or more highly virulent diseases into vulnerable populations of native mammals, eventually causing extinctions. The extinction was biased toward larger-sized species because smaller species have greater resilience because of their life history traits (e.g., shorter gestation time, greater population sizes, etc.). Humans are thought to be the cause because other earlier immigrations of mammals into North America from Eurasia did not cause extinctions.
Diseases imported by people have been responsible for extinctions in the recent past; for example, bringing avian malaria to Hawaii has had a major impact on the isolated birds of the island.
If a disease was indeed responsible for the end-Pleistocene extinctions, then there are several criteria it must satisfy (see Table 7.3 in MacPhee & Marx 1997). First, the pathogen must have a stable carrier state in a reservoir species. That is, it must be able to sustain itself in the environment when there are no susceptible hosts available to infect. Second, the pathogen must have a high infection rate, such that it is able to infect virtually all individuals of all ages and sexes encountered. Third, it must be extremely lethal, with a mortality rate of c. 50–75%. Finally, it must have the ability to infect multiple host species without posing a serious threat to humans. Humans may be infected, but the disease must not be highly lethal or able to cause an epidemic.
One suggestion is that pathogens were transmitted by the expanding humans via the domesticated dogs they brought with them. Unfortunately for such a theory it can not account for several major extinction events, notably Australia and North America. Dogs did not arrive in Australia until approximately 35,000 years after the first humans arrived and approximately 30,000 years after the megafaunal extinction was complete and as such can not be implicated. In contrast numerous species including wolves, mammoths, camelids and horses had emigrated continually between Asia and North America over the past 100,000 years. For the disease hypothesis to be applicable in the case of the Americas it would require that the population remain immunologically naive despite this constant transmission of genetic and pathogenic material.
Arguments against the hyperdisease hypothesis
Scenario
The Second-Order Predation Hypothesis says that as humans entered the New World they continued their policy of killing predators, which had been successful in the Old World but because they were more efficient and because the fauna, both herbivores and carnivores, were more naive, they killed off enough carnivores to upset the ecological balance of the continent, causing overpopulation, environmental exhaustion, and environmental collapse. The hypothesis accounts for changes in animal, plant, and human populations.
The scenario is as follows:
Support
This has been supported by a computer model, the Pleistocene Extinction Model (PEM), which, using the same assumptions and values for all variables (herbivore population, herbivore recruitment rates, food needed per human, herbivore hunting rates, etc.) other than those for hunting of predators. It compares the Overkill hypothesis (predator hunting = 0) with Second-Order Predation (predator hunting varied between 0.01 and 0.05 for different runs). The findings are that Second Order-Predation is more consistent with extinction than is Overkill (results graph at left).
The PEM is the only test of multiple hypotheses and is the only model to specifically test combination hypotheses by artificially introducing sufficient climate change to cause extinction. When Overkill and Climate Change are combined they balance each other out. Climate Change reduces the number of plants, Overkill removes animals, therefore fewer plants are eaten. Second-Order Predation combined with Climate Change exacerbates the effect of Climate Change. (results graph at right).
The second-order predation hypothesis is supported by the observation above that there was a massive increase in bison populations.
Second-order predation and other theories
Arguments against the second-order predation hypothesis
Arguments against the second-order predation plus climate hypothesis
Comet hypothesis
First publicly presented at the Spring 2007 joint assembly of the American Geophysical Union in Acapulco, Mexico, the comet hypothesis suggests that the mass extinction was caused by a swarm of comets 12,900 years ago. Using photomicrograph analysis, research published in January 2009 has found evidence of nanodiamonds in the soil from six sites across North America including Arizona, Minnesota, Oklahoma, South Carolina and two Canadian sites. Similar research found nanodiamonds in the Greenland ice sheet.
Arguments against the comet hypothesis
Debate around this hypothesis has included, among other things, the lack of an impact crater, relatively small increased level of iridium in the soil, and the relative probability of such an event.