
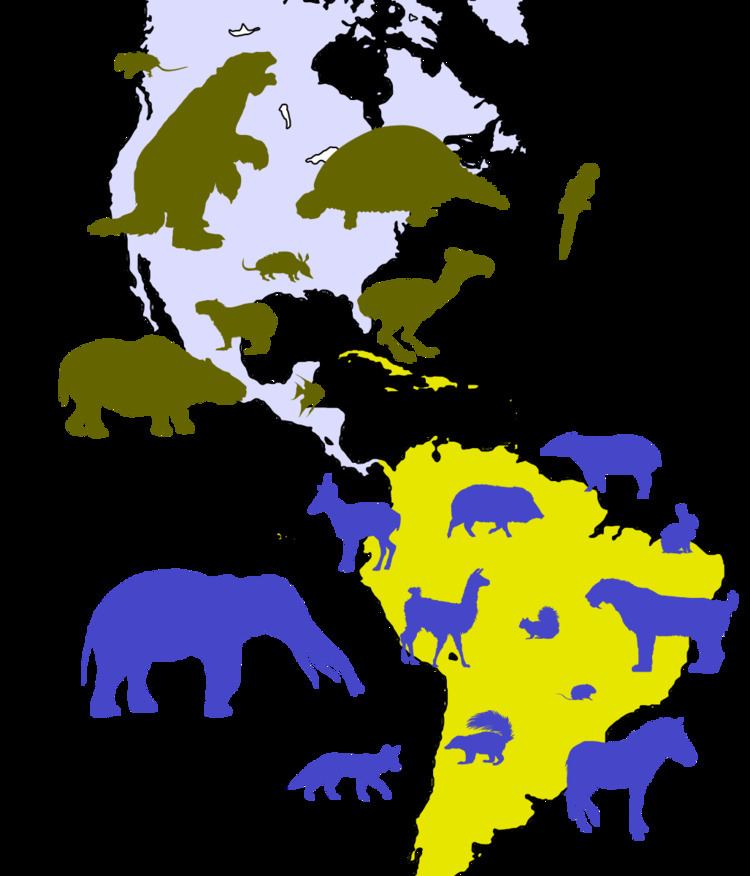
 | ||
The Great American Interchange was an important paleozoogeographic event in which land and freshwater fauna migrated from North America via Central America to South America and vice versa, as the volcanic Isthmus of Panama rose up from the sea floor and bridged the formerly separated continents. The migration peaked dramatically around three million years (Ma) ago during the Piacenzian age.
Contents
- South Americas endemic fauna
- Island hopping waif dispersers
- The Great American Biotic Interchange
- Reasons for success or failure
- Late Pleistocene extinctions
- South American invasions of North America exclusive of Central America
- South American invasions that only extended to Central America
- North American invasions of South America
- References
It resulted in the joining of the Neotropic (roughly South America) and Nearctic (roughly North America) ecozones definitively to form the Americas. The interchange is visible from observation of both biostratigraphy and nature (neontology). Its most dramatic effect is on the zoogeography of mammals but it also gave an opportunity for reptiles, amphibians, arthropods, weak-flying or flightless birds, and even freshwater fish to migrate.
The occurrence of the interchange was first discussed in 1876 by the "father of biogeography", Alfred Russel Wallace. Wallace had spent 1848–1852 exploring and collecting specimens in the Amazon basin. Others who made significant contributions to understanding the event in the century that followed include Florentino Ameghino, W. D. Matthew, W. B. Scott, Bryan Patterson, George Gaylord Simpson and S. David Webb.
Analogous interchanges occurred earlier in the Cenozoic, when the formerly isolated land masses of India and Africa made contact with Eurasia c. 50 and 30 Ma ago, respectively.
South America's endemic fauna
After the late Mesozoic breakup of Gondwana, South America spent most of the Cenozoic era as an island continent whose "splendid isolation" allowed its fauna to evolve into many forms found nowhere else on Earth, most of which are now extinct. Its endemic mammals initially consisted primarily of metatherians (marsupials and sparassodonts), xenarthrans, and a diverse group of native ungulates: notoungulates (the "southern ungulates"), litopterns, astrapotheres (e.g. Trigonostylops, Astrapotherium), and pyrotheres (e.g. Pyrotherium). A few non-therian mammals – monotremes, gondwanatheres, dryolestids and possibly cimolodont multituberculates – were also present in the Paleocene; while none of these diversified significantly and most lineages did not survive long, forms like Necrolestes and Patagonia remained as recently as the Miocene.
Marsupials appear to have traveled via Gondwanan land connections from South America through Antarctica to Australia in the late Cretaceous or early Tertiary. One living South American marsupial, the monito del monte, has been shown to be more closely related to Australian marsupials than to other South American marsupials; however, it is the most basal australidelphian, meaning that this superorder arose in South America and then colonized Australia after the monito del monte split off. A 61-Ma-old platypus-like monotreme fossil from Patagonia may represent an Australian immigrant. Paleognath birds (ratites and South American tinamous) may have migrated by this route around the same time, more likely in the direction from South America to Australia/New Zealand. Other taxa that may have dispersed by the same route (if not by flying or floating across the ocean) are parrots, chelid turtles and (extinct) meiolaniid turtles.
Marsupials present in South America included didelphimorphs (opossums) and several other small groups; larger predatory relatives of these also existed, like the borhyaenids and the sabertooth Thylacosmilus (sparassodont metatherians which are no longer considered to be true marsupials).
Metatherians (and a few xenarthran armadillos like Macroeuphractus) were the only South American mammals to specialize as carnivores; their relative inefficiency created openings for nonmammalian predators to play more prominent roles than usual (similar to the situation in Australia). Sparassodonts shared the ecological niches for large predators with fearsome flightless "terror birds" (phorusrhacids), whose closest extant relatives are the seriemas. Through the skies over late Miocene South America (6 Ma ago) soared the largest flying bird known, the teratorn Argentavis, with a wing span of 6 m or more, which may have subsisted in part on the leftovers of Thylacosmilus kills. Terrestrial ziphodont sebecid crocodilians were also present at least through the middle Miocene and maybe to the Miocene-Pliocene boundary. Some of South America's aquatic crocodilians, such as Gryposuchus, Mourasuchus and Purussaurus, reached monstrous sizes, with lengths up to 12 m (comparable to the largest Mesozoic crocodyliforms). They shared their habitat with one of the largest turtles of all time, the 3.3 m (11 ft) Stupendemys.
Xenarthrans are a curious group of mammals that developed morphological adaptations for specialized diets very early in their history. In addition to those extant today (armadillos, anteaters and tree sloths), a great diversity of larger types were present, including pampatheres, the ankylosaur-like glyptodonts, predatory euphractines, various ground sloths, some of which reached the size of elephants (e.g. Megatherium), and even semiaquatic to aquatic marine sloths.
The notoungulates and litopterns had many strange forms, like Macrauchenia, a camel-like litoptern with a small proboscis. They also produced a number of familiar-looking body types that represent examples of parallel or convergent evolution: one-toed Thoatherium had legs like those of a horse, Pachyrukhos resembled a rabbit, Homalodotherium was a semi-bipedal clawed browser like a chalicothere, and horned Trigodon looked like a rhino. Both groups started evolving in the Lower Paleocene, possibly from condylarth stock, diversified, dwindled before the great interchange, and went extinct at the end of the Pleistocene. The pyrotheres and astrapotheres were also strange but were less diverse and disappeared earlier, well before the interchange.
The North American fauna was a typical boreoeutherian one (supplemented with Afrotherian proboscids).
Island-hopping 'waif dispersers'
The invasions of South America started about 40 Ma ago (middle Eocene), when caviomorph rodents arrived in South America. Their subsequent vigorous diversification displaced some of South America's small marsupials and gave rise to – among others – capybaras, chinchillas, viscachas, and New World porcupines. (The independent development of spines by New and Old World porcupines is another example of parallel evolution.) This invasion most likely came from Africa. The crossing from West Africa to the northeast corner of Brazil was much shorter then, due to continental drift, and may have been aided by island hopping (e.g. via St. Paul's Rocks, if they were an inhabitable island at the time) and westward oceanic currents. Crossings of the ocean were accomplished when at least one fertilised female (more commonly a group of animals) accidentally floated over on driftwood or mangrove rafts. (Island-hopping caviomorphs would subsequently colonize the West Indies as far as the Bahamas, reaching the Greater Antilles by the early Oligocene.) Over time, some caviomorph rodents evolved into larger forms that competed with some of the native South American ungulates, which may have contributed to the gradual loss of diversity suffered by the latter after the early Oligocene.
Later (by 36 Ma ago) primates followed, again from Africa in a fashion similar to that of the rodents. Primates capable of migrating had to be small. Like caviomorph rodents, South American monkeys are believed to be a clade (i.e., monophyletic). However, although they would have had little effective competition, all extant New World monkeys appear to derive from a radiation that occurred long afterwards, in the Early Miocene about 18 Ma ago. Subsequent to this, monkeys apparently most closely related to titis island-hopped to Cuba, Hispaniola and Jamaica. Additionally, a find of seven 21-Ma-old apparent cebid teeth in Panama suggests that South American monkeys had dispersed across the seaway separating Central and South America by that early date. However, all extant Central American monkeys are believed to be descended from much later migrants, and there is as yet no evidence that these early Central American cebids established an extensive or long-lasting population, perhaps due to a shortage of suitable rainforest habitat at the time.
Remarkably, the descendents of those few bedraggled waifs that crawled ashore from their rafts of African flotsam in the Eocene now constitute more than twice as many of South America's species as the descendents of all the nonflying mammals previously resident on the continent (372 caviomorph and monkey species versus 136 marsupial and xenarthran species).
Many of South America's bats may have arrived from Africa during roughly the same period, possibly with the aid of intervening islands, although by flying rather than floating. Noctilionoid bats ancestral to those in the neotropical families Furipteridae, Mormoopidae, Noctilionidae, Phyllostomidae, and Thyropteridae are thought to have reached South America from Africa in the Eocene, possibly via Antarctica. Similarly, molossid bats may have reached South America from Africa in as many as five dispersals, starting in the Eocene. Emballonurid bats may have also reached South America from Africa about 30 Ma ago, based on molecular evidence. Vespertilionid bats may have arrived in five dispersals from North America and one from Africa. Natalid bats are thought to have arrived during the Pliocene from North America via the Caribbean.
Tortoises also arrived in South America in the Oligocene. It was long thought that they had come from North America, but a recent comparative genetic analysis concludes that the South American genus Chelonoidis (formerly part of Geochelone) is actually most closely related to African hingeback tortoises. Tortoises are aided in oceanic dispersal by their ability to float with their heads up, and to survive up to six months without food or water. South American tortoises then went on to colonize the West Indies and Galápagos Islands. A number of clades of American geckos seem to have rafted over from Africa during both the Paleogene and Neogene. Skinks of the related genera Mabuya and Trachylepis apparently dispersed across the Atlantic from Africa to South America and Fernando de Noronha, respectively, during the last 9 Ma. Surprisingly, South America's burrowing amphisbaenians and blind snakes also appear to have rafted from Africa, as does the hoatzin, a weak-flying bird of South American rainforests.
The earliest traditionally recognized mammalian arrival from North America was a procyonid that island-hopped from Central America before a land bridge formed, around 7.3 Ma ago. This was South America's first eutherian carnivore. South American procyonids then diversified into forms now extinct (e.g. the "dog-coati" Cyonasua, which evolved into the bear-like Chapalmalania). However, all extant procyonid genera appear to have originated in North America. It has been suggested that the first South American procyonids may have contributed to the extinction of sebecid crocodilians by eating their eggs, but this view has not been universally viewed as plausible. The procyonids were followed to South America by rafting/island-hopping hog-nosed skunks and sigmodontine rodents The oryzomyine tribe of sigmodontine rodents went on to colonize the Lesser Antilles up to Anguilla.
One group has proposed that a number of large Neartic herbivores actually reached South America as early as 9–10 Ma ago, in the late Miocene, via the "Baudo pathway", an early land bridge that was probably incomplete and required some swimming/island-hopping to traverse. The limited evidence for these early immigrants may reflect their presence primarily in the Amazon basin, an area where fewer fossils have been collected. These taxa are: a proboscidean (Amahuacatherium), peccaries (Sylvochoerus and Waldochoerus), tapirs and a palaeomerycid (from a family probably ancestral to cervids), Surameryx; the paleomerycids were probably unable to successfully colonize South America.
Similarly, megalonychid and mylodontid ground sloths island-hopped to North America by 9 Ma ago. Megalonychids had colonized the Antilles previously, by the early Miocene. (Megatheriid and nothrotheriid ground sloths did not migrate north until the formation of the isthmus.) Terror birds may have also island-hopped to North America as early as 5 Ma ago.
The Caribbean islands were populated primarily by species from South America. This was due to the prevailing direction of oceanic currents, rather than to a competition between North and South American forms. (Except in the case of Jamaica, oryzomyine rodents of North American origin were able to enter the region only after invading South America.)
The Great American Biotic Interchange
The formation of the Isthmus of Panama led to the last and most conspicuous wave, the great interchange, around 3 Ma ago. This included the immigration of North American ungulates (including camelids, tapirs, deer and horses), proboscids (gomphotheres), carnivorans (including felids like cougars and saber-toothed cats, canids, mustelids, procyonids and bears) and a number of types of rodents into South America. The larger members of the reverse migration, besides ground sloths and terror birds, were glyptodonts, pampatheres, capybaras and the notoungulate Mixotoxodon (the only South American ungulate known to have invaded Central America).
In general, the initial net migration was symmetrical. Later on, however, the Neotropic species proved far less successful than the Nearctic. This misfortune happened both ways. Northwardly migrating animals often were not able to compete for resources as well as the North American species already occupying the same ecological niches; those that did become established were not able to diversify much. Southwardly migrating Nearctic species established themselves in larger numbers and diversified considerably more, and are thought to have caused the extinction of a large proportion of the South American fauna. (There were no extinctions in North America plainly linked to South American immigrants.) Although terror birds were able to invade part of North America, their success was temporary; this lineage disappeared about two million years ago. The other large warm-blooded Neotropic predators fared no better. Native South American ungulates also did poorly, with only several of the largest forms, Macrauchenia and a few toxodontids, withstanding the northern onslaught. (Among the notoungulates, the mesotheriids and hegetotheriids did manage to hold on until the Pleistocene.) On the other hand, South America's small marsupials survived in large numbers, while the primitive-looking xenarthrans proved to be surprisingly competitive and became the most successful invaders of North America. The African immigrants, the caviomorph rodents and platyrrhine monkeys, were less impacted by the interchange than most of South America's 'old-timers', although the caviomorphs suffered a significant loss of diversity, including the elimination of the largest forms (e.g. the dinomyids). With the exception of the North American porcupine and several extinct porcupines and capybaras, however, they did not migrate past Central America.
The initial wave of southwardly migrating Nearctic carnivorans rapidly occupied the South American predatory niches, displacing phorusrhacids and sparassodonts, as well as eliminating Chapalmalania. It has been argued that canids probably played the major role in the borhyaenids' extinction; they are ecologically and morphologically more similar to them than other carnivorans, and are also the most diverse family of modern carnivorans on the continent. The paucity of early competition and plentiful prey seems to have allowed short-faced bears to rapidly evolve into the largest known bear or terrestrial mammalian carnivore species; Arctotherium angustidens is estimated to have weighed around 1600 kg. Later species of Arctotherium exhibited a trend towards smaller size and a more omnivorous diet, probably due to increasing competition from later-arriving or evolving carnivores. In contrast, Smilodon showed a trend toward increasing body size that culminated in the appearance of S. populator, at up to nearly 500 kg the most massive felid known.
Due in large part to the success of the xenarthrans, one area of South American ecospace the Nearctic invaders were unable to dominate was the niches for megaherbivores. Before 12,000 years ago, South America was home to about 25 species of herbivores weighing more than 1000 kg, consisting of Neotropic ground sloths, glyptodonts and toxodontids, as well as gomphotheres and camelids of Nearctic origin. Native South American forms made up about 75% of these species. However, none of these megaherbivores have survived.
Armadillos, opossums and porcupines are present in North America today because of the Great American Interchange. Opossums and porcupines were among most successful northward migrants, reaching as far as Canada and Alaska, respectively. Most major groups of xenarthrans were present in North America up until the end-Pleistocene Quaternary extinction event (as a result of at least eight successful invasions of temperate North America, and at least six more invasions of Central America only). Among the megafauna, ground sloths were notably successful emigrants; Megalonyx spread as far north as the Yukon and Alaska, and might well have eventually reached Eurasia if the extinction event had not intervened.
Generally speaking, however, the dispersal and subsequent explosive adaptive radiation of sigmodontine rodents throughout South America (leading to over 80 currently recognized genera) was vastly more successful (both spatially and by number of species) than any northward migration of South American mammals. Other examples of North American mammal groups that diversified conspicuously in South America include canids and cervids, both of which currently have 3 or 4 genera in North America, 2 or 3 in Central America, and 6 in South America. Although Canis currently ranges only as far south as Panama, South America still has more extant canid genera than any other continent.
The effect of formation of the isthmus on the marine biota of the area was the inverse of its effect on terrestrial organisms, a development that has been termed the "Great American Schism". The connection between the east Pacific Ocean and the Caribbean (the Central American Seaway) was severed, setting now-separated populations on divergent evolutionary paths. Caribbean species also had to adapt to an environment of lower productivity after the inflow of nutrient-rich water of deep Pacific origin was blocked.
Reasons for success or failure
The eventual triumph of the Nearctic migrants was ultimately based on geography, which played into the hands of the northern invaders in two crucial respects. The first was a matter of climate. Any species that reached Panama from either direction obviously had to be able to tolerate moist tropical conditions. Those migrating southward would then be able to occupy much of South America without encountering climates that were markedly different. However, northward migrants would have encountered drier and/or cooler conditions by the time they reached the vicinity of the Trans-Mexican Volcanic Belt. The challenge this climatic asymmetry (see map on right) presented was particularly acute for Neotropic species specialized for tropical rainforest environments, who had little prospect of penetrating beyond Central America. As a result, Central America currently has 41 mammal species of Neotropical origin, compared to only 3 for temperate North America. However, species of South American origin (marsupials, xenarthrans, caviomorph rodents and monkeys) still comprise only 21% of species from nonflying, nonmarine mammal groups in Central America, while North American invaders constitute 49% of species from such groups in South America. Thus, climate alone cannot fully account for the greater success of species of Nearctic origin during the interchange.
The second and more important advantage geography gave to the northerners is related to the land area available for their ancestors to evolve in. During the Cenozoic, North America was periodically connected to Eurasia via Beringia, allowing multiple migrations back and forth to unite the faunas of the two continents. Eurasia was connected in turn to Africa, which contributed further to the species that made their way to North America. South America, on the other hand, was connected only to Antarctica and Australia, two much smaller and less hospitable continents, and only in the early Cenozoic. Moreover, this land connection does not seem to have carried much traffic (apparently no mammals other than marsupials and perhaps a few monotremes ever migrated by this route), particularly in the direction of South America. This means that Northern Hemisphere species arose over a land area roughly six times greater than was available to South American species. North American species were thus products of a larger and more competitive arena, where evolution would have proceeded more rapidly. They tended to be more efficient and brainier, generally able to outrun and outwit their South American counterparts, who were products of an evolutionary backwater. These advantages can be clearly seen in the cases of ungulates and their predators, where South American forms were replaced wholesale by the invaders.
The greater eventual success of South America's African immigrants compared to its native early Cenozoic mammal fauna is another example of this phenomenon, since the former evolved over a greater land area; their ancestors migrated from Eurasia to Africa, two significantly larger continents, before finding their way to South America.
Against this backdrop, the ability of South America's xenarthrans to compete effectively against the northerners represents a special case. The explanation for the xenarthrans' success lies in part in their idiosyncratic approach to defending against predation, based on possession of body armor and/or formidable claws. The xenarthrans did not need to be fleet-footed or quick-witted to survive. Such a strategy may have been forced on them by their low metabolic rate (the lowest among the therians). Their low metabolic rate may in turn have been advantageous in allowing them to subsist on less abundant and/or less nutritious food sources. Unfortunately, the defensive adaptations of the large xenarthrans would have offered little protection against humans armed with spears and other projectiles.
Late Pleistocene extinctions
At the end of the Pleistocene epoch, about 12,000 years ago, three dramatic developments occurred in the Americas at roughly the same time (geologically speaking). Paleoindians invaded and occupied the New World, the last glacial period came to an end, and a large fraction of the megafauna of both North and South America went extinct. This wave of extinctions swept off the face of the Earth many of the successful participants of the Great American Interchange, as well as other species that had not migrated. All the pampatheres, glyptodonts, ground sloths, equids, proboscids, dire wolves, lions and Smilodon species of both continents disappeared. The last of the South and Central American notoungulates and litopterns died out, as well as North America's giant beavers, dholes, native cheetahs, scimitar cats, and many of its antilocaprid, bovid, cervid, tapirid and tayassuid ungulates. Some groups disappeared over most or all of their original range but survived in their adopted homes, e.g. South American tapirs, camelids and tremarctine bears (cougars and jaguars may have been temporarily reduced to South American refugia also). Others, such as capybaras, survived in their original range but died out in areas they had migrated to. Notably, this extinction pulse eliminated all Neotropic migrants to North America larger than about 15 kg (the size of a big porcupine), and all native South American mammals larger than about 65 kg (the size of a big capybara or giant anteater). In contrast, the largest surviving native North American mammal, the wood bison, can exceed 900 kg, and the largest surviving Nearctic migrant to South America, Baird's tapir, can reach 400 kg.
The near-simultaneity of the megafaunal extinctions with the glacial retreat and the peopling of the Americas has led to proposals that both climate change and human hunting played a role. Although the subject is contentious, a number of considerations suggest that human activities were pivotal. The extinctions did not occur selectively in the climatic zones that would have been most affected by the warming trend, and there is no plausible general climate-based megafauna-killing mechanism that could explain the continent-wide extinctions. The climate change took place worldwide, but had little effect on the megafauna in areas like Africa and southern Asia, where megafaunal species had coevolved with humans. Numerous very similar glacial retreats had occurred previously within the ice age of the last several Ma without ever producing comparable waves of extinction in the Americas or anywhere else. Similar megafaunal extinctions have occurred on other recently populated land masses (e.g. Australia, Japan, Madagascar, New Zealand, and many smaller islands around the world, such as Cyprus, Crete, Tilos and New Caledonia) at different times that correspond closely to the first arrival of humans at each location. These extinction pulses invariably swept rapidly over the full extent of a contiguous land mass, regardless of whether it was an island or a hemisphere-spanning set of connected continents. This was true despite the fact that all the larger land masses involved (as well as many of the smaller ones) contained multiple climatic zones that would have been affected differently by any climate changes ongoing at the time. However, on sizable islands far enough offshore from newly occupied territory to escape immediate human colonization, megafaunal species sometimes survived for many thousands of years after they or related species became extinct on the mainland; examples include giant kangaroos in Tasmania, giant Chelonoidis tortoises of the Galápagos Islands (formerly also of South America), giant Dipsochelys tortoises of the Seychelles (formerly also of Madagascar), giant meiolaniid turtles on Lord Howe Island, New Caledonia and Vanuatu (previously also of Australia), ground sloths on the Antilles, Steller's sea cows off the Commander Islands and woolly mammoths on Wrangel Island and Saint Paul Island. The glacial retreat may have played a primarily indirect role in the extinctions in the Americas by simply facilitating the movement of humans southeastward from Beringia down to North America. The reason that a number of groups went extinct in North America but lived on in South America (while there are no examples of the opposite pattern) appears to be that the dense rainforest of the Amazon basin and the high peaks of the Andes provided environments that afforded a degree of protection from human predation.
South American invasions of North America exclusive of Central America
Extant or extinct (†) North American taxa whose ancestors migrated out of South America:
South American invasions that only extended to Central America
Extant or extinct (†) Central American taxa whose ancestors migrated out of South America:
North American invasions of South America
Extant or extinct (†) South American taxa whose ancestors migrated out of North America (considered as including Central America):