
Clade Metatheria Higher classification Metatheria | Phylum Chordata Scientific name Marsupialia Rank Infraclass | |
 | ||
Lower classifications Opossum, Wombat, Macropods, Diprotodontia, Dasyurids | ||
Marsupial 18 kinds of marsupials
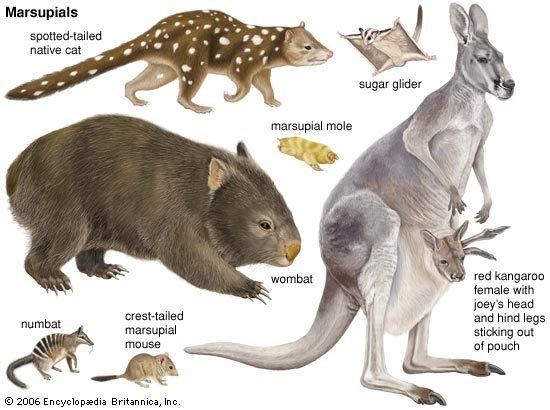
Marsupials. are any members of the mammalian infraclass Marsupialia. All extant marsupials are endemic to Australasia and the Americas. A distinctive characteristic common to these species is that most of the young are carried in a pouch. Well-known marsupials include kangaroos, wallabies, koalas, possums, opossums, wombats, Tasmanian devils, and the recently extinct thylacines. Others include the numbat, the bandicoot, the bettong, the bilby, the quoll, and the quokka.
Contents
- Marsupial 18 kinds of marsupials
- Marsupials for kids marsupial mammals marsupial animals in australia tanzania and americas
- Taxonomy
- Anatomy
- Skull and teeth
- Torso
- General and convergences
- Reproductive system
- Male reproductive system
- Female reproductive system
- Evolution
- References

Marsupials represent the clade originating from the last common ancestor of extant metatherians. Like other mammals in the Metatheria, they give birth to relatively undeveloped young that often reside with the mother in a pouch, for a certain amount of time. Close to 70% of the 334 extant species occur on the Australian continent (the mainland, Tasmania, New Guinea and nearby islands). The remaining 100 are found in the Americas — primarily in South America, but thirteen in Central America, and one in North America, north of Mexico.

Marsupials for kids marsupial mammals marsupial animals in australia tanzania and americas
Taxonomy
Taxonomically, the two primary divisions of Marsupialia are: American and Australian marsupials. The order Microbiotheria (which has only one species, the monito del monte) is found in South America, but is believed to be more closely related to Australian marsupials. There are many small arboreal species in each group. The term "opossum" is used to refer to American species (though "possum" is a common diminutive), while similar Australian species are properly called "possums". Again, shrew opossums are more closely related to australidelphians than to true opossums.
†Extinct

Cladogram based on Gallus, S. et al. (2015)
Anatomy

Marsupials have the typical characteristics of mammals—e.g., mammary glands, three middle ear bones, and true hair. There are, however, striking differences as well as a number anatomical features that separate them from Eutherians.
In addition to the front pouch, which contains multiple nipples for the protection and sustenance of their young, marsupials have other common structural features. Ossified patellae are absent in most modern marsupials (though a small number of exceptions are reported) and epipubic bones are present. Marsupials (and monotremes) also lack a gross communication (corpus callosum) between the right and left brain hemispheres.
Skull and teeth
The skull has peculiarities in comparison to higher mammals. In general, the skull is relatively small and tight. Holes (foramen lacrimale) are located in the front of the orbit. The cheekbone is enlarged and extends further to the rear. The angular extension (processus angularis) of the lower jaw is bent toward the center. Another feature is the hard palate which, in contrast to the higher mammals' foramina, always have more openings. The teeth differ from that of placental mammals, so that all taxa except wombats have a different number of incisors in the upper and lower jaws. The early marsupials had a dental formula from 5 / 4-1 / 1-3 / 3-4 / 4, that is, per pine half; they have five maxilla or four mandibular incisors, one canine, three premolars and four molars, for a total of 50 teeth. Some taxa, such as the opossum, have the original number of teeth. In other groups the number of teeth is reduced. Marsupials in many cases have 40 to 50 teeth, significantly more than placental mammals. The upper jaw has a high number of incisors, up to ten, and they have more molars than premolars. The second set of teeth grows in only at the 3rd premolar: all remaining teeth are already created as permanent teeth.
Torso
Few general characteristics describe their skeleton. In addition to details in the construction of the ankle, bones (Ossa epubica) are characteristic, two from the pubic bone of the pelvis, which is a forwardly projecting bone. Since these are present in males and pouchless species, it is believed that they originally had nothing to do with reproduction, but served in the muscular approach to the movement of the hind limbs. The egg-laying platypus have marsupial bones. This could be explained by an original feature of mammals. Marsupial reproductive organs differ from the higher mammals. For them, the reproductive tract is doubled. The females have two uteri and two vaginas, and before birth, a birth canal forms between them, the pseudo-vagina. The males have a split or double penis lying in front of the scrotum.
A pouch is present in some but not all species. Some marsupials have a permanent bag, whereas in others the pouch develops during gestation, as with the shrew opossum, where the young are hidden only by skin folds or in the fur of the mother. The arrangement of the pouch is variable to allow the offspring to receive maximum protection. Locomotive kangaroos have a pouch opening at the front, while many others that walk or climb on all fours have the opening in the back. Usually, only females have a pouch, but the male water opossum has a pouch that is used to accommodate his genitalia while swimming or running.
General and convergences
Marsupials have adapted to many habitats, reflected in the wide variety in their build. The largest living marsupial, the red kangaroo, grows up to 1.8 metres (5.9 ft) in height and 90 kilograms (200 lb) in weight, but extinct genera, such as Diprotodon, were significantly larger and heavier. The smallest members of this group are the marsupial mice, which often reach only 5 centimetres (2.0 in) in body length.
Some species resemble higher mammals and are examples of convergent evolution. The extinct Thylacine strongly resembled the placental wolf, hence its nickname "Tasmanian wolf". Flying and the associated ability to glide occurred both with marsupials (as with sugar gliders) and some higher mammals (as with flying squirrels), which developed independently. Other groups such as the kangaroo, however, do not have placental counterparts.
Reproductive system
Marsupials' reproductive systems differ markedly from those of placental mammals. The female develops a kind of yolk sac in her womb which delivers nutrients to the embryo. Embryos of some marsupials additionally form placenta-like organs that connect them to the uterine wall, although it is not certain that they transfer nutrients from the mother to the embryo. Pregnancy is very short, typically 4 to 5 weeks, and the embryo is born at a very young stage of development.
The evolution of reproduction in marsupials, and speculation about the ancestral state of mammalian reproduction, have engaged discussion since the end of the 19th century. Both sexes possess a cloaca, which is connected to a urogenital sac used to store waste before expulsion. The bladder of marsupials functions as a site to concentrate urine and empties into the common urogenital sinus in both females and males.
Male reproductive system
Most male marsupials, except for macropods and marsupial moles, have a bifurcated penis, separated into two columns, so that the penis has two ends corresponding to the females' two vaginas. The penis is used only for inseminating females, and is separate from the urinary tract. It curves forward when erect, and when not erect, it is retracted into the body in an S-shaped curve. Neither marsupials nor monotremes possess a baculum. The shape of the glans penis varies among marsupial species.
The male thylacine had a pouch that acted as a protective sheath, covering his external reproductive organs while he ran through thick brush.
The shape of the urethral grooves of the males' genitalia is used to distinguish between Monodelphis brevicaudata, Monodelphis domestica, and Monodelphis americana. The grooves form 2 separate channels that form the ventral and dorsal folds of the erectile tissue. Several species of dasyurid marsupials can also be distinguished by their penis morphology.
The prostate is proportionally larger in marsupials than in placental mammals. During the breeding season, the male tammar wallaby's prostate and bulbourethral gland enlarge. However, there does not appear to be any seasonal difference in the weight of the testes.
Female reproductive system
Female marsupials have two lateral vaginas, which lead to separate uteri, but both open externally through the same orifice. A third canal, the median vagina, is used for birth. This canal can be transitory or permanent. The definitive placenta in all marsupials is generated by the yolk sac. Among three fetal membranes in mammals, the yolk sac, allantois, and amnion, only the first two form a placenta. The evolution of placentation in vertebrates is linked to the evolution of viviparity, a reproductive system in which the females retain their eggs to give birth to their young. Marsupials give birth at a very early stage of development (about four to five weeks); after birth, newborn marsupials crawl up the bodies of their mothers and attach themselves to a nipple, which is located on the underside of the mother either inside a pouch called the marsupium or open to the environment. To crawl to the nipple and attach to it, the marsupial must have well-developed forelimbs and facial structures. This is accomplished by accelerating forelimb and facial development in marsupials compared to placental mammals, which results in decelerated development of such structures as the hindlimb and brain. There they remain for a number of weeks, attached to the nipple. The offspring are eventually able to leave the marsupium for short periods, returning to it for warmth, protection, and nourishment.
Early development
An early birth removes a developing marsupial from its mother's body much sooner than in placental mammals, thus marsupials have not developed a complex placenta to protect the embryo from its mother's immune system. Though early birth puts the tiny newborn marsupial at a greater environmental risk, it significantly reduces the dangers associated with long pregnancies, as there is no need to carry a large fetus to full term in bad seasons. Marsupials are extremely altricial animals, needing to be intensely cared for immediately following birth (cf. precocial).
Because newborn marsupials must climb up to their mother's nipples, their front limbs are much more developed than the rest of their bodies at the time of birth. This requirement has been argued to have resulted in the limited range of locomotor adaptations in marsupials compared to placentals. Marsupials must develop grasping forepaws during their early youth, making the transition from these limbs into hooves, wings, or flippers, as some groups of placental mammals have done, far more difficult. However, several marsupials do possess atypical forelimb morphologies, such as the hooved forelimbs of the pig-footed bandicoot, suggesting that the range of forelimb speciation is a lot less limited than usually assumed.
An infant marsupial is known as a joey. Marsupials have a very short gestation period (about four to five weeks), and the joey is born in an essentially fetal state. The blind, furless, miniature newborn, the size of a jelly bean, crawls across its mother's fur to make its way into the pouch, where it latches onto a teat for food. It will not re-emerge for several months, during which time it develops fully. After this period, the joey begins to spend increasing lengths of time out of the pouch, feeding and learning survival skills. However, it returns to the pouch to sleep, and if danger threatens, it will seek refuge in its mother's pouch for safety.
Joeys stay in the pouch for up to a year in some species, or until the next joey is born. A marsupial joey is unable to regulate its own body temperature and relies upon an external heat source. Until the joey is well-furred and old enough to leave the pouch, a pouch temperature of 30–32 °C (86–90 °F) must be constantly maintained.
Joeys are born with "oral shields". In species without pouches or with rudimentary pouches these are more developed than in forms with well-developed pouches, implying a role in maintaining the young attached to the mother's nipple.
Evolution
The relationships among the three extant divisions of mammals (monotremes, marsupials, and placentals) were long a matter of debate among taxonomists. Most morphological evidence comparing traits such as number and arrangement of teeth and structure of the reproductive and waste elimination systems as well as most genetic and molecular evidence favors a closer evolutionary relationship between the marsupials and placental mammals than either has with the monotremes.
The ancestors of marsupials, part of a larger group called metatherians, probably split from those of placental mammals (eutherians) during the mid-Jurassic period, though no fossil evidence of metatherians themselves are known from this time. Fossil metatherians are distinguished from eutherians by the form of their teeth; metatherians possess four pairs of molar teeth in each jaw, whereas eutherian mammals (including true placentals) never have more than three pairs. Using this criterion, the earliest known metatherian is Sinodelphys szalayi, which lived in China around 125 million years ago (mya). This makes it a contemporary to some early eutherian species which have been found in the same area.
The oldest metatherian fossils are found in present-day China. About 100 mya, the supercontinent Pangaea was in the process of splitting into the northern continent Laurasia and the southern continent Gondwana, with what would become China and Australia already separated by the Tethys Ocean. From there, metatherians spread westward into modern North America (still attached to Eurasia), where the earliest true marsupials are found. Marsupials are difficult to distinguish from other fossils, as they are characterized by aspects of the reproductive system which do not normally fossilize (including pouches) and by subtle changes in the bone and tooth structure that show a metatherian is part of the marsupial crown group (the most exclusive group that contains all living marsupials). The earliest definite marsupial fossil belongs to the species Peradectes minor, from the Paleocene of Montana, dated to about 65 million years ago. From their point of origin in Laurasia, marsupials spread to South America, which was connected to North America until around 65 mya. Laurasian marsupials eventually died off, for not entirely clear reasons; convention has it that they disappeared due to competition with placentals, but this is no longer accepted to be the primary reason.
Marsurpials, Peradectes and the related Herpetotheriidae are nested within a clade of metatherians that also included a variety of Cretaceous North American taxa.
In South America, the opossums evolved and developed a strong presence, and the Paleogene also saw the evolution of shrew opossums (Paucituberculata) alongside non-marsupial metatherian predators such as the borhyaenids and the saber-toothed Thylacosmilus. South American niches for mammalian carnivores were dominated by these marsupial and sparassodont metatherians. While placental predators were absent, the metatherians did have to contend with avian (terror bird) and terrestrial crocodylomorph competition. South America and Antarctica remained connected until 35 mya, as shown by the unique fossils found there. North and South America were disconnected until about three million years ago, when the Isthmus of Panama formed. This led to the Great American Interchange. Sparassodonts disappeared for unclear reasons – again, this has classically assumed as competition from carnivoran placentals, but the last sparassodonts co-existed with a few small carnivorans like procyonids and canines, and disappeared long before the arrival of macropredatory forms like felines, while didelphimorphs (opossums) invaded Central America, with the Virginia opossum reaching as far north as Canada.
Marsupials reached Australia via Antarctica about 50 mya, shortly after Australia had split off. This suggests a single dispersion event of just one species, most likely a relative to South America's monito del monte (a microbiothere, the only New World australidelphian). This progenitor may have rafted across the widening, but still narrow, gap between Australia and Antarctica. In Australia, they radiated into the wide variety seen today. Modern marsupials appear to have reached the islands of Borneo and Sulawesi relatively recently via Australia. A 2010 analysis of retroposon insertion sites in the nuclear DNA of a variety of marsupials has confirmed all living marsupials have South American ancestors. The branching sequence of marsupial orders indicated by the study puts Didelphimorphia in the most basal position, followed by Paucituberculata, then Microbiotheria, and ending with the radiation of Australian marsupials. This indicates that Australidelphia arose in South America, and reached Australia after Microbiotheria split off.
In Australia, terrestrial placental mammals disappeared early in the Cenozoic (their most recent known fossils being 55 million-year-old teeth resembling those of condylarths) for reasons that are not clear, allowing marsupials to dominate the Australian ecosystem. Extant native Australian terrestrial placental mammals (such as hopping mice) are relatively recent immigrants, arriving via island hopping from Southeast Asia.
Genetic analysis suggests a divergence date between the marsupials and the placentals at 160 million years ago. The ancestral number of chromosomes has been estimated to be 2n = 14.
A new hypothesis suggests that South American microbiotheres resulted from a back-dispersal from eastern Gondwana due to new cranial and post-cranial marsupial fossils from the Djarthia murgonensis from the early Eocene Tingamarra Local Fauna in Australia that indicate the Djarthia murgonensis is the most plesiomorphic, the oldest unequivocal australidelphian, and may be the ancestral morphotype of the Australian marsupial radiation.