
Latin systema respiratorium FMA 7158 | TA 06.0.00.000 | |
 | ||
The respiratory system (called also respiratory apparatus, ventilatory system) is a biological system consisting of specific organs and structures used for the process of respiration in an organism. The respiratory system is involved in the intake and exchange of oxygen and carbon dioxide between an organism and the environment.
Contents
- Horses
- Elephants
- Birds
- Reptiles
- Amphibians
- Fish
- Arthropods
- Insects
- Molluscs
- Ventilation
- Control
- Mechanics of Inhalation and exhalation
- Gas exchange
- Physiological responses to low atmospheric pressures
- Immune functions
- Metabolic and endocrine functions of the lungs
- Vocalization
- Temperature control
- Coughing and sneezing
- Humans and mammals
- Disease
- Plants
- References
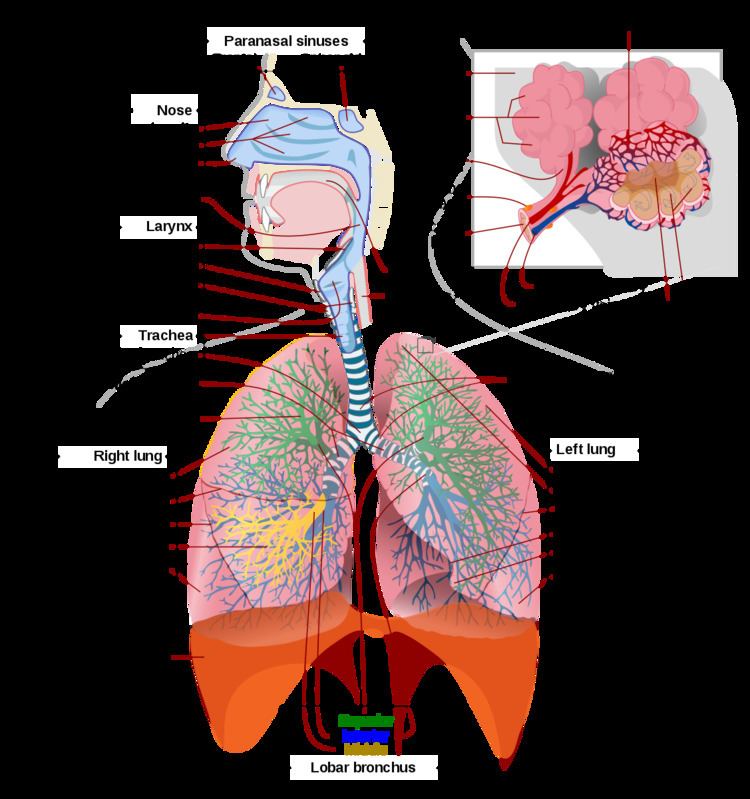
In air-breathing vertebrates like human beings, respiration takes place in the respiratory organs called lungs. The passage of air into the lungs to supply the body with oxygen is known as inhalation, and the passage of air out of the lungs to expel carbon dioxide is known as exhalation; this process is collectively called breathing or ventilation. In humans and other mammals, the anatomical features of the respiratory system include trachea, bronchi, bronchioles, lungs, and diaphragm. Molecules of oxygen and carbon dioxide are passively exchanged, by diffusion, between the gaseous external environment and the blood. This exchange process occurs in the alveoli (air sacs) in the lungs.
In fish and many invertebrates, respiration takes place through the gills. Other animals, such as insects, have respiratory systems with very simple anatomical features, and in amphibians even the skin plays a vital role in gas exchange. Plants also have respiratory systems but the directionality of gas exchange can be opposite to that in animals. The respiratory system in plants also includes anatomical features such as holes on the undersides of leaves known as stomata.
Horses
Horses are obligate nasal breathers which means that they are different from many other mammals because they do not have the option of breathing through their mouths and must take in oxygen through their noses.
Elephants
The elephant is the only animal known to have no pleural space. Rather, the parietal and visceral pleura are both composed of dense connective tissue and joined to each other via loose connective tissue. This lack of a pleural space, along with an unusually thick diaphragm, are thought to be evolutionary adaptations allowing the elephant to remain underwater for long periods of time while breathing through its trunk which emerges as a snorkel.
Birds
The respiratory system of birds differs significantly from that found in mammals, containing unique anatomical features such as air sacs. The lungs of birds also do not have the capacity to inflate as birds lack a diaphragm and a pleural cavity. Gas exchange in birds occurs between air capillaries and blood capillaries, rather than in alveoli.
Reptiles
The anatomical structure of the lungs is less complex in reptiles than in mammals, with reptiles lacking the very extensive airway tree structure found in mammalian lungs. Gas exchange in reptiles still occurs in alveoli however, reptiles do not possess a diaphragm. Thus, breathing occurs via a change in the volume of the body cavity which is controlled by contraction of intercostal muscles in all reptiles except turtles. In turtles, contraction of specific pairs of flank muscles governs inspiration or expiration.
Amphibians
Both the lungs and the skin serve as respiratory organs in amphibians. The ventilation of the lungs in amphibians uses positive pressure ventilation. Muscles lower the floor of the oral cavity, enlarging it and drawing in air through the nostrils (which uses the same mechanics - pressure, volume, and diffusion - as a mammalian lung). With the nostrils and mouth closed, the floor of the oral cavity is forced up, which forces air down the trachea into the lungs.The skin of these animals is highly vascularized and moist, with moisture maintained via secretion of mucus from specialized cells. While the lungs are of primary importance to breathing control, the skin's unique properties aid rapid gas exchange when amphibians are submerged in oxygen-rich water.
Fish
In most fish, respiration takes place through gills. (See also aquatic respiration.) Lungfish, however, do possess one or two lungs. The labyrinth fish have developed a special organ that allows them to take advantage of the oxygen of the air.
Arthropods
Some species of crab use a respiratory organ called a branchiostegal lung. Its gill tissue is formed so as to increase the surface area and the lung is more suited to taking oxygen from the air than from water. Some of the smallest spiders and mites can breathe simply by exchanging gas through the surface of the body. Larger spiders, scorpions and other arthropods use a primitive book lung.
Insects
Most insects breath passively through their spiracles (special openings in the exoskeleton) and the air reaches the body by means of a series of smaller and smaller pipes called 'trachaea' when their diameter is relatively large and 'tracheoles' when their diameter is very small. Diffusion of gases is effective over small distances but not over larger ones, this is one of the reasons insects are all relatively small. Insects which do not have spiracles and trachaea, such as some Collembola, breathe directly through their skins, also by diffusion of gases. The number of spiracles an insect has is variable between species, however they always come in pairs, one on each side of the body, and usually one per segment. Some of the Diplura have eleven, with four pairs on the thorax, but in most of the ancient forms of insects, such as Dragonflies and Grasshoppers there are two thoracic and eight abdominal spiracles. However, in most of the remaining insects there are less. It is at this level of the tracheoles that oxygen is delivered to the cells for respiration. The trachea are water-filled due to the permeable membrane of the surrounding tissues. During exercise, the water level retracts due to the increase in concentration of lactic acid in the muscle cells. This lowers the water potential and the water is drawn back into the cells via osmosis and air is brought closer to the muscle cells. The diffusion pathway is then reduced and gases can be transferred more easily.
Insects were once believed to exchange gases with the environment continuously by the simple diffusion of gases into the tracheal system. More recently, however, large variation in insect ventilatory patterns have been documented and insect respiration appears to be highly variable. Some small insects do demonstrate continuous respiration and may lack muscular control of the spiracles. Others, however, utilize muscular contraction of the abdomen along with coordinated spiracle contraction and relaxation to generate cyclical gas exchange patterns and to reduce water loss into the atmosphere. The most extreme form of these patterns is termed discontinuous gas exchange cycles (DGC).
Molluscs
Molluscs generally possess gills that allow exchange of oxygen from an aqueous environment into the circulatory system. These animals also possess a heart that pumps blood which contains hemocyaninine as its oxygen-capturing molecule. Hence, this respiratory system is similar to that of vertebrate fish. The respiratory system of gastropods can include either gills or a lung.
Ventilation
In respiratory physiology, the ventilation rate is the rate at which gas enters or leaves the lung. It is categorized under the following definitions:
Control
Ventilation occurs via the respiratory center in the medulla oblongata and the pons of the brainstem. These areas of the brain form a series of interconnected brain cells within the lower and middle brain stem which receive information about the arterial partial pressure of oxygen (
The breathing rate increases when the
Mechanics of Inhalation and exhalation
In mammals, breathing in (inhalation) at rest is primarily due to the contraction and flattening of the diaphragm, a domed muscle that separates the thoracic cavity from the abdominal cavity. When the diaphragm contracts it pushes the abdominal organs downward, but since the pelvic floor prevents the lowermost abdominal organs from moving in that direction, the abdomen, in fact, bulges forwards (or outwards). In the process the size of the thoracic cavity has increased in volume (as has the volume of the body as a whole). This increased thoracic volume results in a fall in pressure in the thorax, which causes the expansion of the lungs. During exhalation (breathing out), at rest, the diaphragm relaxes, returning the chest and abdomen to a position which is determined by their anatomical elasticity (i.e. the position in the cadaver, or in an animal that has been given a muscle relaxant under anesthesia). This is the "resting mid-position" of the thorax when the lungs contain the functional residual capacity of air, which in the adult human has a volume of about 2.5 liters. Resting exhalation lasts about twice as long as inhalation because the diaphragm relaxes more gently than it contracts during inhalation. This prevents undue narrowing of the airways, from which the air escapes more easily than from the alveoli.
During heavy breathing (hyperventilation), as, for instance, during exercise, the "accessory muscles of inhalation" (of which the first to be recruited are the intercostal muscles, but include a large number of other muscles - see below) pull the ribs upwards, both in the front and on the sides. This increases the volume of the rib cage, adding to the volume increase caused by the descending diaphragm. During the ensuing exhalation the rib cage is actively pulled downwards (front and sides) by the abdominal muscles, which not only decreases the size of the rib cage, but also pushes the abdominal organs upwards against the diaphragm which consequently bulges deeply into the thorax. The end-exhalatory lung volume is now well below the resting mid-position and contains far less air than the resting "functional residual capacity". However, in a normal mammal, the lungs cannot be emptied completely. In an adult human there is always still at least 1 liter of residual air left in the lungs after maximum exhalation.
The entirely unconscious and automatic breathing on which the life of the animal depends can be temporarily over-ridden by conscious or emotion-driven movements of air in and out of the lungs. Speech in humans is generated by a specialized form of exhalation, but other forms of communication (e.g. crying, yelping, yawning, barking, baying, hissing, panting, sighing, shouting, laughing etc.) also rely on a balance between breathing for blood gas homeostasis and the emotional or other messages that need to be conveyed to the animal's conspecifics.
Ten muscles can be used for inhalation:
Diaphragm, Intercostal Muscles, Scalenes, Pectoralis Minor, Serratus Anterior, Sternocleidomastoid, Levator Costarum, Upper / Superior Trapezius, Latissimus Dorsi, and Subclavius.
Eight are used for forced exhalation:
Internal intercostal, Obliquus Internus, Obliquus Externus, Levator Ani, Triangularis Sterni, Transversalis, Pyramidalis, and Rectus Abdominus.
Gas exchange
The major function of the pulmonary gas exchanger in mammals is the equilibration of the blood gases with those in the alveolar air. The alveolar and pulmonary capillary gases equilibrate across the blood–air barrier, a membrane which forms the walls of the pulmonary alveoli, consisting of the alveolar epithelial cells, their basement membranes and the endothelial cells of the pulmonary capillaries. This blood gas barrier is extremely thin (in humans, on average, 2.2 μm thick) but is extremely strong. This strength comes from the type IV collagen in between the endothelial and epithelial cells. Damage can occur to this barrier at a pressure difference of around 5.3 kPa (40 mmHg). This membrane is folded into about 300 million small air sacs called alveoli (each between 75 and 300 µm in diameter) branching off from the bronchioles in the lungs, thus providing an extremely large surface area (approximately 145 m2) for gas exchange to occur.
The lungs of an average person at rest, and breathing normally contain between about 2.5 and 3 liters of alveolar air. This semi-stagnant volume of air that always remains in the lung alveoli after a normal exhalation is termed the functional residual capacity. With each breath only about 350 ml (i.e. less than 15%) of this alveolar air is expelled into the ambient air to be replaced with the same volume of fresh, but moistened, atmospheric air. It is therefore obvious that the composition of the alveolar air (or functional residual capacity) changes very little under normal circumstances: the alveolar partial pressure of oxygen (
This marked difference between the composition of the alveolar air and that of the ambient air can be maintained because the functional residual capacity is contained in dead-end sacs connected to the outside air by fairly narrow and relatively long tubes (the airways: nose, pharynx, larynx, trachea, bronchi and their branches down to the bronchioles). This anatomy and the fact that the lungs are not emptied and re-inflated with each breath, provides the animal with a very special portable atmosphere, whose composition differs significantly from the present-day ambient air. It is this portable atmosphere (the functional residual capacity) to which the blood and therefore the body tissues are exposed - not to the outside air.
All the blood returning from the body tissues to the right side of the heart flows through the pulmonary capillaries before being pumped around the body again. On its passage through the lungs the blood comes into close contact with the alveolar air, separated from it by a very thin diffusion membrane which is only, on average, about 2 μm thick. The gas pressures in the blood will therefore rapidly equilibrate with those in the alveoli, ensuring that the arterial blood that circulates to all the tissues throughout the body has an oxygen tension of 14 kPa (105 mmHg), and a carbon dioxide tension of 5.3 kPa (40 mmHg). These arterial partial pressures of oxygen and carbon dioxide are homeostatically controlled. A rise in the arterial
Since the blood arriving in the pulmonary capillaries has a
It is only as a result of accurately maintaining the composition of the 3 liters alveolar air that with each breath some carbon dioxide is discharged into the atmosphere and some oxygen is taken up from the outside air. If more carbon dioxide than usual has been lost by a short period of hyperventilation, respiration will be slowed down or halted until the alveolar
If these homeostats are compromised, then a respiratory acidosis, or a respiratory alkalosis will occur. In the long run these can be compensated by renal adjustments to the H+ and HCO3− concentrations in the plasma; but since this takes time, the hyperventilation syndrome can, for instance, occur when agitation or anxiety cause a person to breathe fast and deeply thus causing a distressing respiratory alkalosis through the blowing off of too much CO2 from the blood into the outside air.
Oxygen has a very low solubility in water, and is therefore carried in the blood loosely combined with hemoglobin. The oxygen is held on the hemoglobin by four ferrous iron-containing heme groups per hemoglobin molecule. When all the heme groups carry one O2 molecule each the blood is said to be “saturated” with oxygen, and no further increase in the
Physiological responses to low atmospheric pressures
The alveoli are open (via the airways) to the atmosphere, with the result that alveolar air pressure is exactly the same as the ambient air pressure at sea level, at altitude, or in any artificial atmosphere (e.g. a diving chamber, or decompression chamber) in which the individual is breathing freely. With expansion of the lungs (through lowering of the diaphragm and expansion of the thoracic cage) the alveolar air now occupies a larger volume, and its pressure falls proportionally, causing air to flow in from the surroundings, through the airways, till the pressure in the alveoli is once again at the ambient air pressure. The reverse obviously happens during exhalation. This process (of inhalation and exhalation) is exactly the same at sea level, as on top of Mt. Everest, or in a diving chamber or decompression chamber.
However, as one rises above sea level the density of the air decreases exponentially, halving approximately with every 5500 m rise in altitude. Since the composition of the atmospheric air is almost constant below 80 km, as a result of the continuous mixing effect of the weather, the concentration of oxygen in the air (mmols O2 per liter of ambient air) decreases at the same rate as the fall in air pressure with altitude. Therefore, in order to breathe in the same amount of oxygen per minute, the person has to inhale a proportionately greater volume of air per minute at altitude than at sea level. This is achieved by breathing deeper and faster (i.e. hyperventilating) than at sea level (see below).
There is, however, a complication that increases the volume of air that needs to be inhaled per minute (respiratory minute volume) to provide the same amount of oxygen to the lungs at altitude as at sea level. During inhalation the air is warmed and saturated with water vapor during its passage through the nose and pharynx. Saturated water vapor pressure is dependent only on temperature. At a body core temperature of 37 °C it is 6.3 kPa (47.0 mmHg), irrespective of any other influences, including altitude. Thus at sea level, where the ambient atmospheric pressure is about 100 kPa, the moistened air that flows into the lungs from the trachea consists of water vapor (6.3 kPa), nitrogen (74.0 kPa), oxygen (19.7 kPa) and trace amounts of carbon dioxide and other gases (a total of 100 kPa). In dry air the partial pressure of oxygen at sea level is 21.0 kPa, compared to the 19.7 kPa of oxygen entering the alveolar air. (The tracheal partial pressure of oxygen (19.7 kPa) is 21% of [100 kPa - 6.3 kPa]). At the summit of Mt. Everest (at an altitude of 8,848 m or 29,029 ft) the total atmospheric pressure is 33.7 kPa, of which 7.1 kPa, or 21%, is oxygen. The air entering the lungs also has a total pressure of 33.7 kPa, of which 6.3 kPa is, unavoidably, water vapor (as it is at sea level). This reduces the partial pressure of oxygen entering the alveoli to 5.8 kPa, or 21% of [33.7 kPa – 6.3 kPa = 27.4 kPa]. The reduction in the partial pressure of oxygen in the inhaled air is therefore substantially greater than the reduction of the total atmospheric pressure at altitude would suggest (on Mt Everest: 5.8 kPa vs. 7.1 kPa).
A further minor complication exists at altitude. Consider the following theoretical scenario. If the volume of the lungs were to be instantaneously doubled at the beginning of inhalation, the air pressure inside the lungs would be halved. This happens regardless of altitude. Thus, halving of the sea level air pressure (100 kPa) results in an intrapulmonary air pressure of 50 kPa. Doing the same at 5500 m, where the atmospheric pressure is only 50 kPa, the intrapulmonary air pressure falls to 25 kPa. Therefore, the same change in lung volume at sea level results in a 50 kPa difference in pressure between the ambient air and the intrapulmonary air, whereas it result in a difference of only 25 kPa at 5500 m. The driving pressure forcing air into the lungs during inhalation is therefore halved at this altitude. The rate of inflow of air into the lungs during inhalation at sea level is therefore twice that which occurs at 5500 m. However, in reality, inhalation and exhalation occur far more gently and less abruptly than in the example given. The differences between the atmospheric and intrapulmonary pressures, driving air in and out of the lungs during the breathing cycle, are in the region of only 2-3 kPa. A doubling or more of these small pressure differences could be achieved by only very minor adjustments to the breathing effort at high altitudes.
All of the above influences of low atmospheric pressures on breathing are accommodated primarily by hyperventilating (or hyperpnea). The exact degree of hyperventilation is determined by the blood gas homeostat, which regulates the partial pressures of oxygen (
There are oxygen sensors in the smaller bronchi and bronchioles. In response to low partial pressures of oxygen in the inhaled air these sensors reflexly cause the pulmonary arterioles to constrict. (This is the exact opposite of the corresponding reflex in the tissues, where low arterial
The kidneys measure the oxygen content (rather than the
Immune functions
Airway epithelial cells can secrete a variety of molecules that aid in the defense of lungs. Secretory immunoglobulins (IgA), collectins (including Surfactant A and D), defensins and other peptides and proteases, reactive oxygen species, and reactive nitrogen species are all generated by airway epithelial cells. These secretions can act directly as antimicrobials to help keep the airway free of infection. Airway epithelial cells also secrete a variety of chemokines and cytokines that recruit the traditional immune cells and others to site of infections.
Most of the respiratory system is lined with mucous membranes that contain mucosal-associated lymphoid tissue, which produces white blood cells such as lymphocytes.
Metabolic and endocrine functions of the lungs
In addition to their functions in gas exchange, the lungs have a number of metabolic functions. They manufacture surfactant for local use, as noted above. They also contain a fibrinolytic system that lyses clots in the pulmonary vessels. They release a variety of substances that enter the systemic arterial blood and they remove other substances from the systemic venous blood that reach them via the pulmonary artery. Prostaglandins are removed from the circulation, but they are also synthesized in the lungs and released into the blood when lung tissue is stretched. The lungs also activate one hormone; the physiologically inactive decapeptide angiotensin I is converted to the pressor, aldosterone-stimulating octapeptide angiotensin II in the pulmonary circulation. The reaction occurs in other tissues as well, but it is particularly prominent in the lungs. Large amounts of the angiotensin-converting enzyme responsible for this activation are located on the surface of the endothelial cells of the pulmonary capillaries. The converting enzyme also inactivates bradykinin. Circulation time through the pulmonary capillaries is less than one second, yet 70% of the angiotensin I reaching the lungs is converted to angiotensin II in a single trip through the capillaries. Four other peptidases have been identified on the surface of the pulmonary endothelial cells.
Vocalization
The movement of gas through the larynx, pharynx and mouth allows humans to speak, or phonate. Vocalization, or singing, in birds occurs via the syrinx, an organ located at the base of the trachea. The vibration of air flowing across the larynx (vocal cords), in humans, and the syrinx, in birds, results in sound. Because of this, gas movement is extremely vital for communication purposes.
Temperature control
Panting in dogs, cats and some other animals provides a means of controlling body temperature. This physiological response is used as a cooling mechanism.
Coughing and sneezing
Irritation of nerves within the nasal passages or airways, can induce a cough reflex and sneezing. These responses cause air to be expelled forcefully from the trachea or nose, respectively. In this manner, irritants caught in the mucus which lines the respiratory tract are expelled or moved to the mouth where they can be swallowed. During coughing, contraction of the smooth muscle narrows the trachea by pulling the ends of the cartilage plates together and by pushing soft tissue out into the lumen. This increases the expired airflow rate to dislodge and remove any irritant particle or mucus.
Humans and mammals
The respiratory system lies dormant in the human fetus during pregnancy. At birth, the respiratory system becomes fully functional upon exposure to air, although some lung development and growth continues throughout childhood. Pre-term birth can lead to infants with under-developed lungs. These lungs show incomplete development of the alveolar type II cells, cells that produce surfactant. The lungs of pre-term infants may not function well because the lack of surfactant leads to increased surface tension within the alveoli. Thus, many alveoli collapse such that no gas exchange can occur within some or most regions of an infant's lungs, a condition termed respiratory distress syndrome. Basic scientific experiments, carried out using cells from chicken lungs, support the potential for using steroids as a means of furthering development of type II alveolar cells. In fact, once a pre-mature birth is threatened, every effort is made to delay the birth, and a series of steroid shots is frequently administered to the mother during this delay in an effort to promote lung growth.
Disease
Disorders of the respiratory system can be classified into four general areas:
Coughing is of major importance, as it is the body's main method to remove dust, mucus, saliva, and other debris from the lungs. Inability to cough can lead to infection. Deep breathing exercises may help keep finer structures of the lungs clear from particulate matter, etc.
The respiratory tract is constantly exposed to microbes due to the extensive surface area, which is why the respiratory system includes many mechanisms to defend itself and prevent pathogens from entering the body.
Disorders of the respiratory system are usually treated internally by a pulmonologist and Respiratory Therapist.
Plants
Plants use carbon dioxide gas in the process of photosynthesis, and exhale oxygen gas as waste. The chemical equation of photosynthesis is 6 CO2 (carbon dioxide) and 6 H2O (water) and that makes 6 O2 (oxygen) and C6H12O6 (glucose). What is not expressed in the chemical equation is the capture of energy from sunlight which occurs. Photosynthesis uses electrons on the carbon atoms as the repository for that energy. Respiration is the opposite of photosynthesis. It reclaims the energy to power chemical reactions in cells. In so doing the carbon atoms and their electrons are combined with oxygen forming a gas which is easily removed from both the cells and the organism. Plants use both processes, photosynthesis to capture the energy and respiration to use it.
Plant respiration is limited by the process of diffusion. Plants take in carbon dioxide through holes on the undersides of their leaves known as stoma or pores. However, most plants require little air. Most plants have relatively few living cells outside of their surface because air (which is required for metabolic content) can penetrate only skin deep. However, most plants are not involved in highly aerobic activities, and thus have no need of these living cells.