
 | ||
Gas exchange is a biological process through which different gases are transferred in opposite directions across a specialized respiratory surface. Gases are constantly required by, and produced as a by-product of, cellular and metabolic reactions, so an efficient system for their exchange is extremely important. It is linked with respiration in animals, and both respiration and photosynthesis in plants.
Contents
In respiration, oxygen (O
2) is required to enter cells, while waste carbon dioxide (CO
2) must be excreted; the opposite is true for photosynthesis, in which CO
2 enters plants and O
2 is released. The exchange of gases essentially occurs as a result of diffusion down a concentration gradient: gas molecules moving from an area of high concentration to low concentration.
Diffusion
Diffusion follows Fick’s Law. It is a passive process (no energy is required) affected by factors such as the surface area available, the distance the gas molecules must diffuse across and the concentration gradient.
Gases must first dissolve in a liquid in order to diffuse across a membrane, so all gas exchange systems require a moist environment.
In single-celled organisms, diffusion can occur straight across the cell membrane; as organisms increase in size, so does the distance gases must travel across. (Their surface area-to-volume ratio also decreases.) Diffusion alone is not efficient enough and specialized respiratory systems are required. This is the case with humans and with fish that have evolved circulatory systems: these are able to transport the gases to and from the respiratory surface and maintain a continuous concentration gradient.
In mammals
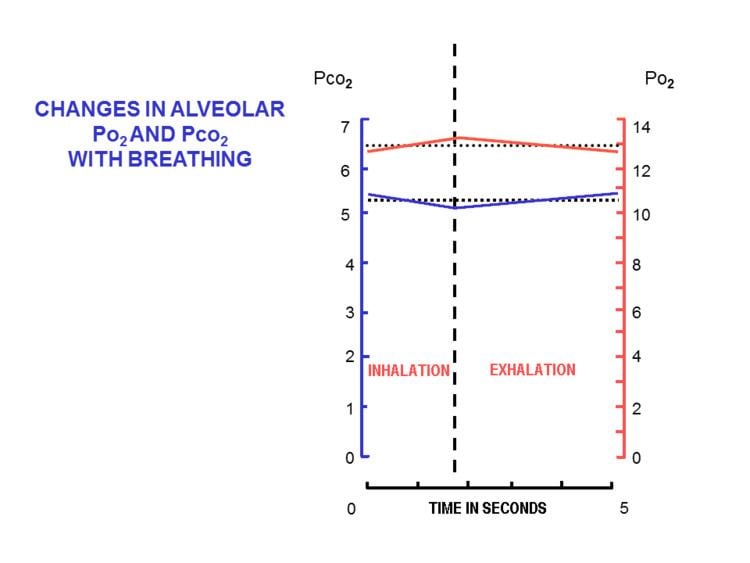
The major function of the pulmonary gas exchanger in mammals is the equilibration of the blood gases with those in the alveolar air. The alveolar and pulmonary capillary gases equilibrate across the blood–air barrier, a membrane which forms the walls of the pulmonary alveoli, consisting of the alveolar epithelial cells, their basement membranes and the endothelial cells of the pulmonary capillaries. This blood gas barrier is extremely thin (in humans, on average, 2.2 μm thick) but is extremely strong. This strength comes from the type IV collagen in between the endothelial and epithelial cells. Damage can occur to this barrier at a pressure difference of around 5.3 kPa (40 mmHg). This membrane is folded into about 300 million small air sacs called alveoli (each between 75 and 300 µm in diameter) branching off from the bronchioles in the lungs, thus providing an extremely large surface area (approximately 145 m2) for gas exchange to occur.
The lungs of an average person at rest, and breathing normally contain between about 2.5 and 3 liters of alveolar air. This semi-stagnant volume of air that always remains in the lung alveoli after a normal exhalation is termed the functional residual capacity. With each breath only about 350 ml (i.e. less than 15%) of this alveolar air is expelled into the ambient air to be replaced with the same volume of fresh, but moistened, atmospheric air. It is therefore obvious that the composition of the alveolar air (or functional residual capacity) changes very little under normal circumstances: the alveolar partial pressure of oxygen (
This marked difference between the composition of the alveolar air and that of the ambient air can be maintained because the functional residual capacity is contained in dead-end sacs connected to the outside air by fairly narrow and relatively long tubes (the airways: nose, pharynx, larynx, trachea, bronchi and their branches down to the bronchioles). This anatomy and the fact that the lungs are not emptied and re-inflated with each breath, provides the animal with a very special portable atmosphere, whose composition differs significantly from the present-day ambient air. It is this portable atmosphere (the functional residual capacity) to which the blood and therefore the body tissues are exposed - not to the outside air.
All the blood returning from the body tissues to the right side of the heart flows through the pulmonary capillaries before being pumped around the body again. On its passage through the lungs the blood comes into close contact with the alveolar air, separated from it by a very thin diffusion membrane which is only, on average, about 2 μm thick. The gas pressures in the blood will therefore rapidly equilibrate with those in the alveoli, ensuring that the arterial blood that circulates to all the tissues throughout the body has an oxygen tension of 14 kPa (105 mmHg), and a carbon dioxide tension of 5.3 kPa (40 mmHg). These arterial partial pressures of oxygen and carbon dioxide are homeostatically controlled. A rise in the arterial
Since the blood arriving in the pulmonary capillaries has a
It is only as a result of accurately maintaining the composition of the 3 liters alveolar air that with each breath some carbon dioxide is discharged into the atmosphere and some oxygen is taken up from the outside air. If more carbon dioxide than usual has been lost by a short period of hyperventilation, respiration will be slowed down or halted until the alveolar
If these homeostats are compromised, then a respiratory acidosis, or a respiratory alkalosis will occur. In the long run these can be compensated by renal adjustments to the H+ and HCO3− concentrations in the plasma; but since this takes time, the hyperventilation syndrome can, for instance, occur when agitation or anxiety cause a person to breathe fast and deeply thus causing a distressing respiratory alkalosis through the blowing off of too much CO2 from the blood into the outside air.
Oxygen has a very low solubility in water, and is therefore carried in the blood loosely combined with hemoglobin. The oxygen is held on the hemoglobin by four ferrous iron-containing heme groups per hemoglobin molecule. When all the heme groups carry one O2 molecule each the blood is said to be “saturated” with oxygen, and no further increase in the
Varying response
During physical exercise, excess carbon dioxide is produced as a result of increased respiration, and muscles and cells require increased oxygen. The body responds to this change by increasing the breathing rate, maximizing the rate of possible gas exchange.
In plants
Gas exchange in plants is dominated by the roles of carbon dioxide and water vapor. CO
2 is the only carbon source for autotrophic organisms, making it essential for the conversion of light into sugar during photosynthesis. Due to the high differences in water potential in the plant versus the surrounding air, water vapor tends to evaporate from plants. Gas exchange is mediated through pores (known as stomata and located mainly on the lower side of leaves) that underlie a complex regulatory system. As the condition of the stomata unavoidably influences both the CO
2 and water vapor exchanges, plants experience a gas exchange dilemma: gaining enough CO
2 without losing too much water.
Gas exchange measurements are common tools in plant science. If the environmental conditions (humidity, CO
2 concentration, light and temperature) are fully controlled, the measurements of CO
2 uptake and water release reveal important information about the CO
2 assimilation and transpiration rates and the intercellular CO
2 concentration, which reveal important information about the photosynthetic condition of the plants.
Oxygen, essential for respiration during the night, plays a minor role in plants' gas exchange as it is always present in sufficient amounts.
In fish
Fish must extract oxygen dissolved in water, not air, which has led to the evolution of gills and opercula. Gills are specialized organs containing filaments and lamellae: the lamellae contain capillaries and provide a large surface area and short diffusion distance, as they are extremely thin.
Water is drawn in through the mouth and passes over the gills in one direction while blood flows through the lamellae in the opposite direction. This countercurrent maintains a steep concentration gradient. Oxygen is able to continually diffuse down its gradient into the blood, and the CO
2 into the water.
Other examples
Insects such as crickets do not have an inner skeleton, so they exchange gases across structures known as trachea and tracheoles: tubes that run directly into the insect's body. Air enters the trachea through spiracles and diffuses into the respiring tissues.
Amphibians are able to use their skin as a respiratory surface. They also have lungs and sometimes gills can be used.