
EC number 4.2.1.1 ExPASy NiceZyme view | CAS number 9001-03-0 | |
 | ||
Carbonic anhydrases
The carbonic anhydrases (or carbonate dehydratases) form a family of enzymes that catalyze the rapid interconversion of carbon dioxide and water to bicarbonate and protons (or vice versa), a reversible reaction that occurs relatively slowly in the absence of a catalyst. The active site of most carbonic anhydrases contains a zinc ion; they are therefore classified as metalloenzymes.
Contents
- Carbonic anhydrases
- Carbonic anhydrase
- Reaction
- Mechanism
- Families
- CA
- Structure and function
- Cadmium containing Carbonic Anhydrase
- Similarities To Other Carbonic Anhydrases
- Differences From Other Carbonic Anhydrases
- Transport of Cadmium
- CDCA like Proteins
- Industrial applications
- References
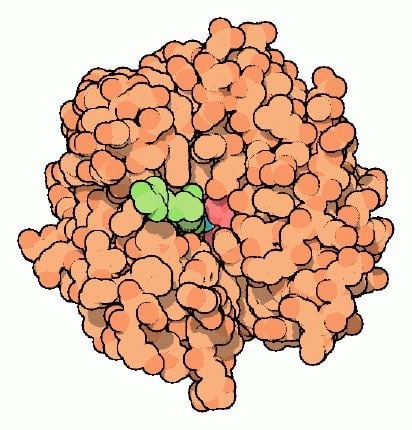
One of the functions of the enzyme in animals is to interconvert carbon dioxide and bicarbonate to maintain acid-base balance in blood and other tissues, and to help transport carbon dioxide out of tissues.
Carbonic anhydrase
Reaction
The reaction catalyzed by carbonic anhydrase is:
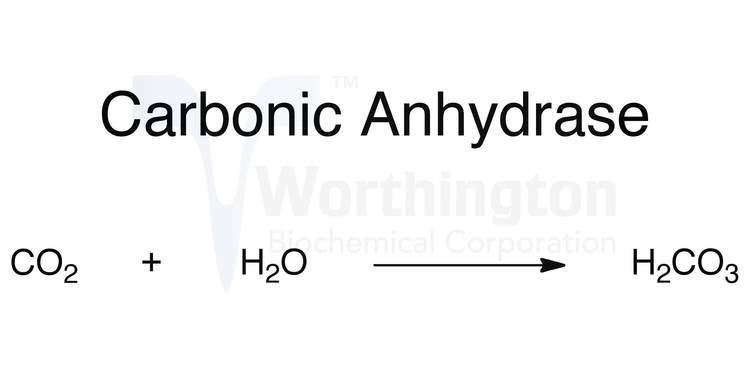
The reaction rate of carbonic anhydrase is one of the fastest of all enzymes, and its rate is typically limited by the diffusion rate of its substrates. Typical catalytic rates of the different forms of this enzyme ranging between 104 and 106 reactions per second.
The reverse reaction is relatively slow (kinetics in the 15-second range) in the absence of a catalyst. This is why a carbonated drink does not instantly degas when opening the container; however it will rapidly degas in the mouth when it comes in contact with carbonic anhydrase that is contained in saliva.
An anhydrase is defined as an enzyme that catalyzes the removal of a water molecule from a compound, and so it is this "reverse" reaction that gives carbonic anhydrase its name, because it removes a water molecule from carbonic acid.
Mechanism
A zinc prosthetic group in the enzyme is coordinated in three positions by histidine side-chains. The fourth coordination position is occupied by water. This causes polarisation of the hydrogen-oxygen bond, making the oxygen slightly more positive, thereby weakening the bond.
A fourth histidine is placed close to the substrate of water and accepts a proton, in an example of general acid - general base catalysis (see the article "Acid catalysis"). This leaves a hydroxide attached to the zinc.
The active site also contains a specificity pocket for carbon dioxide, bringing it close to the hydroxide group. This allows the electron-rich hydroxide to attack the carbon dioxide, forming bicarbonate.
Families
There are at least five distinct CA families (α, β, γ, δ and ζ). These families have no significant amino acid sequence similarity and in most cases are thought to be an example of convergent evolution. The α-CAs are found in humans.
α-CA
Vertebrates, algae and some bacteria have this family of CAs.
The CA enzymes found in mammals are divided into four broad subgroups, which, in turn consist of several isoforms:
There are three additional "acatalytic" CA isoforms (CA-VIII, CA-X, and CA-XI) (CA8, CA10, CA11) whose functions remain unclear.
β-CA
Most prokaryotic and plant chloroplast CAs belong to the beta family. Two signature patterns for this family have been identified:
γ-CA
The gamma class of CAs come from methanogens, methane-producing bacteria that grow in hot springs.
δ-CA
The delta class of CAs has been described in diatoms. The distinction of this class of CA has recently come into question, however.
ζ-CA
The zeta class of CAs occurs exclusively in bacteria in a few chemolithotrophs and marine cyanobacteria that contain cso-carboxysomes. Recent 3-dimensional analyses suggest that ζ-CA bears some structural resemblance to β-CA, particularly near the metal ion site. Thus, the two forms may be distantly related, even though the underlying amino acid sequence has since diverged considerably.
η-CA
The eta family of CAs was recently found in organisms of the genus Plasmodium. These are a group of enzymes previously thought to belong to the alpha family of CAs, however it has been demonstrated that η-CAs have unique features, such as their metal ion coordination pattern.
Structure and function
Several forms of carbonic anhydrase occur in nature. In the best-studied α-carbonic anhydrase form present in animals, the zinc ion is coordinated by the imidazole rings of 3 histidine residues, His94, His96, and His119.
The primary function of the enzyme in animals is to interconvert carbon dioxide and bicarbonate to maintain acid-base balance in blood and other tissues, and to help transport carbon dioxide out of tissues.
There are at least 14 different isoforms in mammals. Plants contain a different form called β-carbonic anhydrase, which, from an evolutionary standpoint, is a distinct enzyme, but participates in the same reaction and also uses a zinc ion in its active site. In plants, carbonic anhydrase helps raise the concentration of CO2 within the chloroplast in order to increase the carboxylation rate of the enzyme RuBisCO. This is the reaction that integrates CO2 into organic carbon sugars during photosynthesis, and can use only the CO2 form of carbon, not carbonic acid or bicarbonate.
Cadmium-containing Carbonic Anhydrase
Marine diatoms have been found to express a new form of ζ carbonic anhydrase. T. weissflogii, a species of phytoplankton common to many marine ecosystems, was found to contain carbonic anhyrdase with a cadmium ion in place of zinc. Previously, it had been believed that cadmium was a toxic metal with no biological function whatsoever. However, this species of phytoplankton appears to have adapted to the low levels of zinc in the ocean by using cadmium when there is not enough zinc. Although the concentration of cadmium in sea water is also low (about 1x10−16 molar), there is an environmental advantage to being able to use either metal depending on which is more available at the time. This type of carbonic anhydrase is therefore cambialistic, meaning it can interchange the metal in its active site with other metals (namely, zinc and cadmium).
Similarities To Other Carbonic Anhydrases
The mechanism of cadmium carbonic anhydrase (CDCA) is essentially the same as that of other carbonic anhydrases in its conversion of carbon dioxide and water into bicarbonate and a proton. Additionally, like the other carbonic anhydrases, CDCA makes the reaction go almost as fast as the diffusion rate of its substrates, and it can be inhibited by sulfonamide and sulfamate derivatives.
Differences From Other Carbonic Anhydrases
Unlike most other carbonic anhydrases, the active site metal ion is not bound by three histidine residues and a hydroxide ion. Instead, it is bound by two cysteine residues, one histidine residue, and a hydroxide ion, which is characteristic of β-CA. Due to the fact that cadmium is a soft acid, it will be more tightly bound by soft base ligands. The sulfur atoms on the cysteine residues are soft bases, thus binding the cadmium more tightly than the nitrogen on histidine residues would. CDCA also has a three-dimensional folding structure that is unlike any other carbonic anhydrase, and its amino acid sequence is dissimilar to the other carbonic anhydrases. It is a monomer with three domains, each one identical in amino acid sequence and each one containing an active site with a metal ion.
Another key difference between CDCA and the other carbonic anhydrases is that CDCA has a mechanism for switching out its cadmium ion for a zinc ion in the event that zinc becomes more available to the phytoplankton than cadmium. The active site of CDCA is essentially "gated" by a chain of nine amino acids with glycine residues at positions 1 and 9. Normally, this gate remains closed and the cadmium ion is trapped inside. However, due to the flexibility and position of the glycine residues, this gate can be opened in order to remove the cadmium ion. A zinc ion can then be put in its place and the gate will close behind it. As a borderline acid, zinc will not bind as tightly to the cysteine ligands as cadmium would, but the enzyme will still be active and reasonably efficient. The metal in the active site can be switched between zinc and cadmium depending on which one is more abundant at the time. It is the ability of CDCA to utilize either cadmium or zinc that likely gives T. weissflogii a survival advantage.
Transport of Cadmium
Cadmium is still considered lethal to phytoplankton in high amounts. Studies have shown that T. weissflogii has an initial toxic response to cadmium when exposed to it. The toxicity of the metal is reduced by the transcription and translation of phytochelatin, which are proteins that can bind and transport cadmium. Once bound by phytochelatin, cadmium is no longer toxic, and it can be safely transported to the CDCA enzyme. It's also been shown that the uptake of cadmium via phytochelatin leads to a significant increase in CDCA expression.
CDCA-like Proteins
Other phytoplankton from different water sources have been tested for the presence of CDCA. It was found that many of them contain proteins that are homologous to the CDCA found in T. weissflogii. This includes species from Great Bay, New Jersey as well as in the Pacific Ocean near the equator. In all species tested, CDCA-like proteins showed high levels of expression even in high concentrations of zinc and in the absence of cadmium. The similarity between these proteins and the CDCA expressed by T. weissflogii varied, but they were always at least 67% similar.
Industrial applications
Modified carbonic anhydrase enzymes have been used to replace methyl diethanolamine ("MDEA") in carbon dioxide capture.