
Latin Diaphragma | Greek διάφραγμα | |
 | ||
Precursor Septum transversum, pleuroperitoneal folds, body wall Artery Pericardiacophrenic artery, musculophrenic artery, inferior phrenic arteries Vein Superior phrenic vein, inferior phrenic vein Nerve Phrenic and lower intercostal nerves | ||
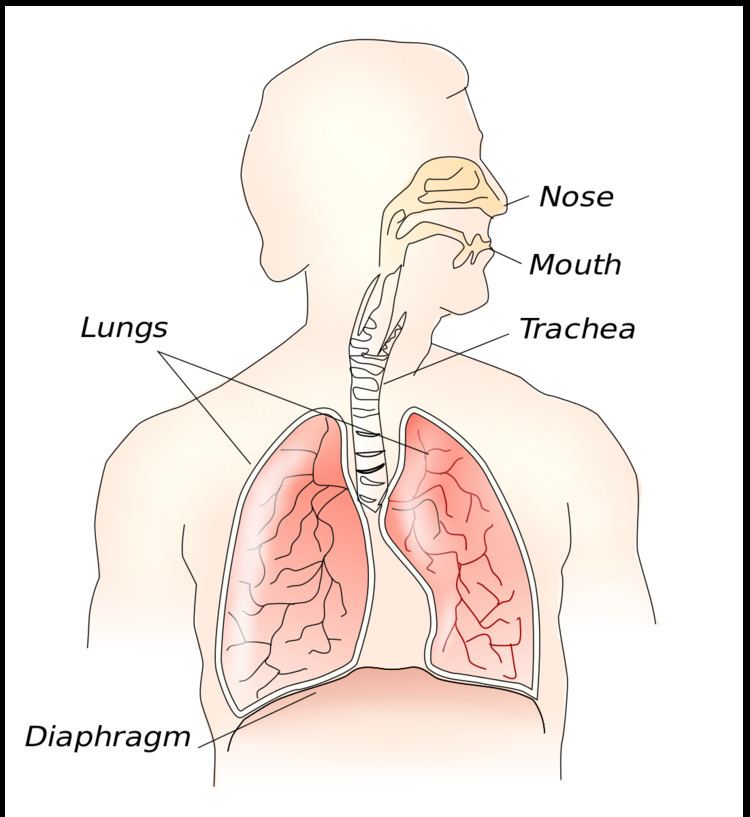
In human anatomy, the thoracic diaphragm, or simply the diaphragm (Ancient Greek: διάφραγμα diáphragma "partition"), is a sheet of internal skeletal muscle that extends across the bottom of the thoracic cavity. The diaphragm separates the thoracic cavity, containing the heart and lungs, from the abdominal cavity and performs an important function in respiration: as the diaphragm contracts, the volume of the thoracic cavity increases and air is drawn into the lungs.
Contents
- Structure
- Crura and central tendon
- Openings
- Innervation
- Blood supply
- Variation
- Development
- Function
- Paralysis
- Herniation
- Imaging
- Other animals
- References
The term diaphragm in anatomy can refer to other flat structures such as the urogenital diaphragm or pelvic diaphragm, but "the diaphragm" generally refers to the thoracic diaphragm. In humans, the diaphragm is slightly asymmetric—its right half is higher up (superior) to the left half, since the large liver rests beneath the right half of the diaphragm.
Other mammals have diaphragms, and other vertebrates such as amphibians and reptiles have diaphragm-like structures, but important details of the anatomy vary, such as the position of the lungs in the abdominal cavity.
Structure
The diaphragm is a C -shaped structure of muscle and fibrous tissue that separates the thoracic cavity from the abdomen. The dome curves upwards. The superior surface of the dome forms the floor of the thoracic cavity, and the inferior surface the roof of the abdominal cavity.
As a dome, the diaphragm has peripheral attachments to structures that make up the abdominal and chest walls. The muscle fibres from these attachments converge in a central tendon, which forms the crest of the dome. Its peripheral part consists of muscular fibers that take origin from the circumference of the inferior thoracic aperture and converge to be inserted into a central tendon.
The muscle fibres of the diaphragm emerge from many surrounding structures. At the front, fibres insert into the xiphoid process and along the costal margin. Laterally, muscle fibers insert into ribs 6-12. In the back, muscle fibres insert into the vertebra at T12 and two appendages, the right and left crus, descend and insert into the lumbar vertebrae at L1 & L2.
There are two lumbocostal arches, a medial and a lateral, on either side.
Crura and central tendon
The left and right crura are tendons that blend of the anterior longitudinal ligament of the vertebral column.
The central tendon of the diaphragm is a thin but strong aponeurosis near the center of the vault formed by the muscle, closer to the front than to the back of the thorax, so that the posterior muscular fibers are the longer.
Openings
There are a number of openings in the diaphragm through which structures pass between the thorax and abdomen. There are three large openings—the aortic, the esophageal, and the caval opening—plus a series of smaller ones.
The inferior vena cava passes through the caval opening, a quadrilateral opening at the junction of the right and middle leaflets of the central tendon, so that its margins are tendinous. Surrounded by tendons, the opening is stretched open every time inspiration occurs. However, there has been argument that the caval opening actually constricts during inspiration. Since thoracic pressure decreases upon inspiration and draws the caval blood upwards toward the right atrium, increasing the size of the opening allows more blood to return to the heart, maximizing the efficacy of lowered thoracic pressure returning blood to the heart. The aorta does not pierce the diaphragm but rather passes behind it in between the left and right crus.
The thoracic spinal levels at which the three major structures pass through the diaphragm can be remembered by the number of letters contained in each structure:
Innervation
The diaphragm is primarily innervated by the phrenic nerve which is formed from the cervical nerves C3, C4 and C5. While the central portion of the diaphragm sends sensory afferents via the phrenic nerve, the peripheral portions of the diaphragm send sensory afferents via the intercostal (T5–T11) and subcostal nerves (T12).
Blood supply
Arteries and veins above and below the diaphragm supply and drain blood.
From above, the diaphragm receives blood from branches of the internal thoracic arteries, namely the pericardiophrenic artery and musculophrenic artery; from the superior phrenic arteries, which arise directly from the thoracic aorta; and from the lower internal intercostal arteries. From below, the inferior phrenic arteries supply the diaphragm.
The diaphragm drains blood into the brachiocephalic veins, azygos veins, and veins that drain into the inferior vena cava and left suprarenal vein.
Variation
The sternal portion of the muscle is sometimes wanting and more rarely defects occur in the lateral part of the central tendon or adjoining muscle fibers.
Development
The thoracic diaphragm develops during embryogenesis, beginning in the third week after fertilization with two processes known as transverse folding and longitudinal folding. The septum transversum, the primitive central tendon of the diaphragm, originates at the rostral pole of the embryo and is relocated during longitudinal folding to the ventral thoracic region. Transverse folding brings the body wall anteriorly to enclose the gut and body cavities. The pleuroperitoneal membrane and body wall myoblasts, from somatic lateral plate mesoderm, meet the septum transversum to close off the pericardio-peritoneal canals on either side of the presumptive esophagus, forming a barrier that separates the peritoneal and pleuropericardial cavities. Furthermore, dorsal mesenchyme surrounding the presumptive esophagus form the muscular crura of the diaphragm.
Because the earliest element of the embryological diaphragm, the septum transversum, forms in the cervical region, the phrenic nerve that innervates the diaphragm originates from the cervical spinal cord (C3,4, and 5). As the septum transversum descends inferiorly, the phrenic nerve follows, accounting for its circuitous route from the upper cervical vertebrae, around the pericardium, finally to innervate the diaphragm.
Function
The diaphragm functions in breathing. During inhalation, the diaphragm contracts and moves in the inferior direction, thus enlarging the volume of the thoracic cavity (the external intercostal muscles also participate in this enlargement). This reduces intra-thoracic pressure: In other words, enlarging the cavity creates suction that draws air into the lungs.
Cavity expansion happens in two extremes, along with intermediary forms. When the lower ribs are stabilized and the central tendon of the diaphragm is mobile, a contraction brings the insertion (central tendon) towards the origins and pushes the lower cavity towards the pelvis, allowing the thoracic cavity to expand downward. This is often called belly breathing. When the central tendon is stabilized and the lower ribs are mobile, a contraction lifts the origins (ribs) up towards the insertion (central tendon) which works in conjunction with other muscles to allow the ribs to slide and the thoracic cavity to expand laterally and upwards.
When the diaphragm relaxes, air is exhaled by elastic recoil of the lung and the tissues lining the thoracic cavity. Assisting this function with muscular effort (called forced exhalation) involves the internal intercostal muscles used in conjunction with the abdominal muscles, which act as an antagonist paired with the diaphragm's contraction.
The diaphragm is also involved in non-respiratory functions, helping to expel vomit, feces, and urine from the body by increasing intra-abdominal pressure, aiding in childbirth, and preventing acid reflux by exerting pressure on the esophagus as it passes through the esophageal hiatus.
In some non-human animals, the diaphragm is not crucial for breathing; a cow, for instance, can survive fairly asymptomatically with diaphragmatic paralysis as long as no massive aerobic metabolic demands are made of it.
Paralysis
If either the phrenic nerve, cervical spine or brainstem is damaged, this will sever the nervous supply to the diaphragm. The most common damage to the phrenic nerve is by bronchial cancer, which usually only affects one side of the diaphragm. Other causes include Guillain–Barré syndrome and systemic lupus erythematosis.
Herniation
A hiatus hernia is a hernia common in adults in which parts of the lower esophagus or stomach that are normally in the abdomen pass bulge abnormally through the diaphragm and are present in the thorax. Hernias are described as rolling, in which the hernia is beside the oesophagus, or sliding, in which the hernia directly involves the esophagus. These hernias are implicated in the development of reflux, as the different pressures between the thorax and abdomen normally act to keep pressure on the esophageal hiatus. With herniation, this pressure is no longer present, and the angle between the cardia of the stomach and the oesophagus disappear. Not all hiatus hernias cause symptoms however, although almost all people with Barrett's oesophagus or oesophagitis have a hiatus hernia.
Hernias may also occur as a result of congenital malformation, a congenital diaphragmatic hernia. When the pleuroperitoneal membranes fail to fuse, the diaphragm does not act as an effective barrier between the abdomen and thorax. Herniation is usually of the left, and commonly through the posterior lumbocostal triangle, although rarely through the anterior foramen of Morgagni. The contents of the abdomen, including the intestines, may be present in the thorax, which may impact development of the growing lungs and lead to hypoplasia. This condition is present in 1 out of 2,000 births. A large herniation has a mortality rate of three out of four, and requires immediate surgical repair.
Imaging
Due to its position separating the thorax and abdomen, fluid abnormally present in the thorax, or air abnormally present in the abdomen, may collect on one side of the diaphragm. An X-ray may reveal this. Pleural effusion, in which there is fluid abnormally present between the two pleurae of the lungs, is detected by an X-ray of the chest, showing fluid collecting in the angle between the ribs and diaphragm. An X-ray may also be used to reveal a pneumoperitoneum, in which there is gas in the abdomen.
An X-ray may also be used to check for herniation.
Other animals
The existence of a membrane separating the pharynx from the stomach can be traced widely among the chordates. Thus the model organism, the marine chordate lancelet, possesses an atriopore by which water exits the pharynx, which has been claimed (and disputed) to be homologous to structures in ascidians and hagfishes. The tunicate epicardium separates digestive organs from the pharynx and heart, but the anus returns to the upper compartment to discharge wastes through an outgoing siphon.
Thus the diaphragm emerges in the context of a body plan that separated an upper feeding compartment from a lower digestive tract, but the point at which it originates is a matter of definition. Structures in fish, amphibians, reptiles, and birds have been called diaphragms, but it has been argued that these structures are not homologous. For instance, the alligator diaphragmaticus muscle does not insert on the esophagus and does not affect pressure of the lower esophageal sphincter. The lungs are located in the abdominal compartment of amphibians and reptiles, so that contraction of the diaphragm expels air from the lungs rather than drawing it into them. In birds and mammals, lungs are located above the diaphragm. The presence of an exceptionally well-preserved fossil of Sinosauropteryx, with lungs located beneath the diaphragm as in crocodiles, has been used to argue that dinosaurs could not have sustained an active warm-blooded physiology, or that birds could not have evolved from dinosaurs. An explanation for this (put forward in 1905), is that lungs originated beneath the diaphragm, but as the demands for respiration increased in warm-blooded birds and mammals, natural selection came to favor the parallel evolution of the herniation of the lungs from the abdominal cavity in both lineages. However, birds do not have diaphragms. They do not breathe in the same way as mammals, and do not rely on creating a negative pressure in the thoracic cavity, at least not to the same extent. They rely on a rocking motion of the keel of the sternum to create local areas of reduced pressure to supply thin, membranous airsacs cranially and caudally to the fixed-volume, non-expansive lungs. A complicated system of valves and air sacs cycles air constantly over the absorption surfaces of the lungs so allowing maximal efficiency of gaseous exchange. Thus, birds do not have the reciprocal tidal breathing flow of mammals. On careful dissection, around eight air sacs can be clearly seen. They extend quite far caudally into the abdomen.