
 | ||
A steroid is an organic compound with four rings arranged in a specific configuration. Examples include the dietary lipid cholesterol, the sex hormones estradiol and testosterone and the anti-inflammatory drug dexamethasone. Steroids have two principal biological functions: certain steroids (such as cholesterol) are important components of cell membranes which alter membrane fluidity, and many steroids are signaling molecules which activate steroid hormone receptors.
Contents
- Nomenclature
- Species distribution and function
- By function
- Intact ring system
- Cleaved contracted and expanded rings
- Biological significance
- Biosynthesis and metabolism
- Mevalonate pathway
- Pharmacological action
- Steroidogenesis
- Alternative pathways
- Catabolism and excretion
- Isolation structure determination and methods of analysis
- Chemical synthesis
- Semisynthesis
- Total synthesis
- Research awards
- References
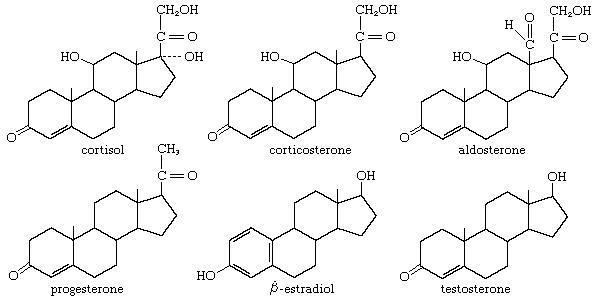
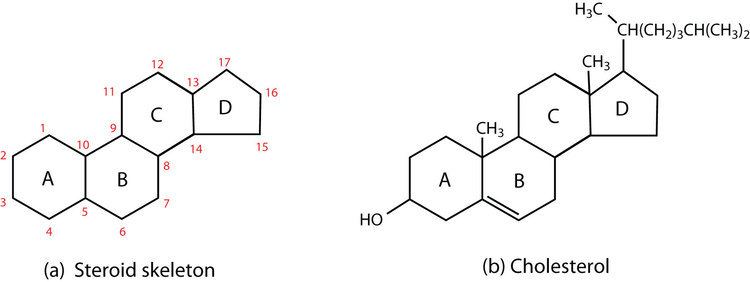
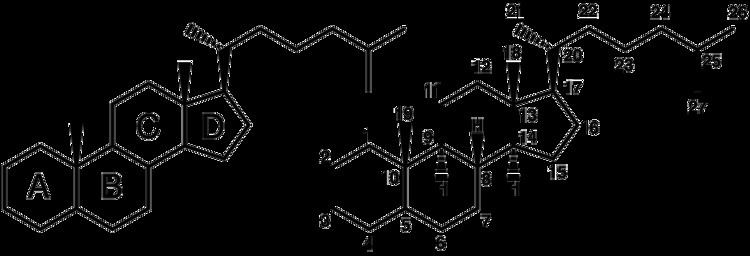
The steroid core structure is composed of seventeen carbon atoms, bonded in four "fused" rings: three six-member cyclohexane rings (rings A, B and C in the first illustration) and one five-member cyclopentane ring (the D ring). Steroids vary by the functional groups attached to this four-ring core and by the oxidation state of the rings. Sterols are forms of steroids with a hydroxyl group at position three and a skeleton derived from cholestane. They can also vary more markedly by changes to the ring structure (for example, ring scissions which produce secosteroids such as vitamin D3).
Hundreds of steroids are found in plants, animals and fungi. All steroids are manufactured in cells from the sterols lanosterol (animals and fungi) or cycloartenol (plants). Lanosterol and cycloartenol are derived from the cyclization of the triterpene squalene.
Nomenclature

Gonane, also known as steran or cyclopentaperhydrophenanthrene, the simplest steroid and the nucleus of all steroids and sterols, is composed of seventeen carbon atoms in carbon-carbon bonds forming four fused rings in a three-dimensional shape. The three cyclohexane rings (A, B, and C in the first illustration) form the skeleton of a perhydro derivative of phenanthrene. The D ring has a cyclopentane structure. When the two methyl groups and eight carbon side chains (at C-17, as shown for cholesterol) are present, the steroid is said to have a cholestane framework. The two common 5α and 5β stereoisomeric forms of steroids exist because of differences in the side of the largely planar ring system where the hydrogen (H) atom at carbon-5 is attached, which results in a change in steroid A-ring conformation.
Examples of steroid structures are:
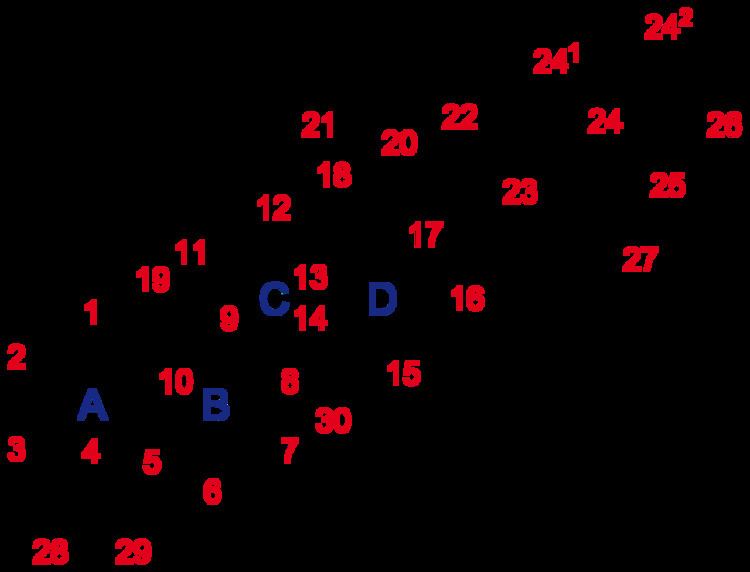
In addition to the ring scissions (cleavages), expansions and contractions (cleavage and reclosing to a larger or smaller rings)—all variations in the carbon-carbon bond framework—steroids can also vary:
For instance, sterols such as cholesterol and lanosterol have an hydroxyl group attached at position C-3, while testosterone and progesterone have a carbonyl (oxo substituent) at C-3; of these, lanosterol alone has two methyl groups at C-4 and cholesterol (with a C-5 to C-6 double bond) differs from testosterone and progesterone (which have a C-4 to C-5 double bond).
Species distribution and function
The following are some common categories of steroids. In eukaryotes, steroids are found in fungi, animals, and plants. Fungal steroids include the ergosterols.
Animal steroids include compounds of vertebrate and insect origin, the latter including ecdysteroids such as ecdysterone (controlling molting in some species). Vertebrate examples include the steroid hormones and cholesterol; the latter is a structural component of cell membranes which helps determine the fluidity of cell membranes and is a principal constituent of plaque (implicated in atherosclerosis). Steroid hormones include:

Plant steroids include steroidal alkaloids found in Solanaceae, the phytosterols, and the brassinosteroids (which include several plant hormones). In prokaryotes, biosynthetic pathways exist for the tetracyclic steroid framework (e.g. in mycobacteria) – where its origin from eukaryotes is conjectured – and the more-common pentacyclic triterpinoid hopanoid framework.
By function
Steroids can be classified functionally. The major classes of steroid hormones, with prominent members and examples of related functions, are:
Intact ring system
Steroids can be classified based on their chemical composition. One example of how MeSH performs this classification is available at the Wikipedia MeSH catalog. Examples of this classification include:
The gonane (steroid nucleus) is the parent 17-carbon tetracyclic hydrocarbon molecule with no alkyl sidechains.
Cleaved, contracted, and expanded rings
Secosteroids (Latin seco, "to cut") are a subclass of steroidal compounds resulting, biosynthetically or conceptually, from scission (cleavage) of parent steroid rings (generally one of the four). Major secosteroid subclasses are defined by the steroid carbon atoms where this scission has taken place. For instance, the prototypical secosteroid cholecalciferol, vitamin D3 (shown), is in the 9,10-secosteroid subclass and derives from the cleavage of carbon atoms C-9 and C-10 of the steroid B-ring; 5,6-secosteroids and 13,14-steroids are similar.
Norsteroids (nor-, L. norma; "normal" in chemistry, indicating carbon removal) and homosteroids (homo-, Greek homos; "same", indicating carbon addition) are structural subclasses of steroids formed from biosynthetic steps. The former involves enzymic ring expansion-contraction reactions, and the latter is accomplished (biomimetically) or (more frequently) through ring closures of acyclic precursors with more (or fewer) ring atoms than the parent steroid framework.
Combinations of these ring alterations are known in nature. For instance, ewes who graze on corn lily ingest cyclopamine (shown) and veratramine, two of a sub-family of steroids where the C- and D-rings are contracted and expanded respectively via a biosynthetic migration of the original C-13 atom. Ingestion of these C-nor-D-homosteroids results in birth defects in lambs: cyclopia from cyclopamine and leg deformity from veratramine. A further C-nor-D-homosteroid (nakiterpiosin) is excreted by Okinawan cyanobacteriosponges – Terpios hoshinota – leading to coral mortality from black coral disease. Nakiterpiosin-type steroids are active against the signaling pathway involving the smoothened and hedgehog proteins, a pathway which is hyperactive in a number of cancers.
Biological significance
Steroids and their metabolites often function as signalling molecules (the most notable examples are steroid hormones), and steroids and phospholipids are components of cell membranes. Steroids such as cholesterol decrease membrane fluidity. Similar to lipids, steroids are highly concentrated energy stores. However, they are not typically sources of energy; in mammals, they are normally metabolized and excreted.
Steroids play critical roles in a number of disorders, including malignancies like prostate cancer, where steroid production inside and outside the tumour promotes cancer cell aggressiveness.
Biosynthesis and metabolism
The hundreds of steroids found in animals, fungi, and plants are made from lanosterol (in animals and fungi; see examples above) or cycloartenol (in plants). Lanosterol and cycloartenol derive from cyclization of the triterpenoid squalene.
Steroid biosynthesis is an anabolic pathway which produces steroids from simple precursors. A unique biosynthetic pathway is followed in animals (compared to many other organisms), making the pathway a common target for antibiotics and other anti-infection drugs. Steroid metabolism in humans is also the target of cholesterol-lowering drugs, such as statins.
In humans and other animals the biosynthesis of steroids follows the mevalonate pathway, which uses acetyl-CoA as building blocks for dimethylallyl pyrophosphate (DMAPP) and isopentenyl pyrophosphate (IPP). In subsequent steps DMAPP and IPP join to form geranyl pyrophosphate (GPP), which synthesizes the steroid lanosterol. Modifications of lanosterol into other steroids are classified as steroidogenesis transformations.
Mevalonate pathway
The mevalonate pathway (also called HMG-CoA reductase pathway) begins with acetyl-CoA and ends with dimethylallyl pyrophosphate (DMAPP) and isopentenyl pyrophosphate (IPP).
DMAPP and IPP donate isoprene units, which are assembled and modified to form terpenes and isoprenoids (a large class of lipids, which include the carotenoids and form the largest class of plant natural products. Here, the isoprene units are joined to make squalene and folded into a set of rings to make lanosterol.
Lanosterol can then be converted into other steroids, such as cholesterol and ergosterol.
Pharmacological action
Two classes of drugs target the mevalonate pathway: statins, which are used to reduce elevated cholesterol levels, and bisphosphonates, which are used to treat a number of bone-degenerative diseases.
Steroidogenesis
Steroidogenesis is the biological process by which steroids are generated from cholesterol and changed into other steroids. The pathways of steroidogenesis differ among species. The major classes of steroid hormones, as noted above (with their prominent members and functions), are the Progestogen, Corticosteroids (corticoids), Androgens, and Estrogens. Human steroidogenesis of these classes occurs in a number of locations:
Alternative pathways
In plants and bacteria, the non-mevalonate pathway uses pyruvate and glyceraldehyde 3-phosphate as substrates.
During diseases pathways otherwise not significant in healthy humans can become utilized. For example, in one form of congenital adrenal hyperplasia an deficiency in the 21-hydroxylase enzymatic pathway leads to an excess of 17α-Hydroxyprogesterone (17-OHP) – this pathological excess of 17-OHP in turn may be converted to dihydrotestosterone (DHT, a potent androgen) through among others 17,20 Lyase (a member of the cytochrome P450 family of enzymes), 5α-Reductase and 3α-Hydroxysteroid dehydrogenase.
Catabolism and excretion
Steroids are primarily oxidized by cytochrome P450 oxidase enzymes, such as CYP3A4. These reactions introduce oxygen into the steroid ring, allowing the cholesterol to be broken up by other enzymes into bile acids. These acids can then be eliminated by secretion from the liver in bile. The expression of the oxidase gene can be upregulated by the steroid sensor PXR when there is a high blood concentration of steroids. Steroid hormones, lacking the side chain of cholesterol and bile acids, are typically hydroxylated at various ring positions or oxidized at the 17 position, conjugated with sulfate or glucuronic acid and excreted in the urine.
Isolation, structure determination, and methods of analysis
Steroid isolation, depending on context, is the isolation of chemical matter required for chemical structure elucidation, derivitzation or degradation chemistry, biological testing, and other research needs (generally milligrams to grams, but often more or the isolation of "analytical quantities" of the substance of interest (where the focus is on identifying and quantifying the substance (for example, in biological tissue or fluid). The amount isolated depends on the analytical method, but is generally less than one microgram. The methods of isolation to achieve the two scales of product are distinct, but include extraction, precipitation, adsorption, chromatography, and crystallization. In both cases, the isolated substance is purified to chemical homogeneity; combined separation and analytical methods, such as LC-MS, are chosen to be "orthogonal"—achieving their separations based on distinct modes of interaction between substance and isolating matrix—to detect a single species in the pure sample. Structure determination refers to the methods to determine the chemical structure of an isolated pure steroid, using an evolving array of chemical and physical methods which have included NMR and small-molecule crystallography. Methods of analysis overlap both of the above areas, emphasizing analytical methods to determining if a steroid is present in a mixture and determining its quantity.
Chemical synthesis
Microbial catabolism of phytosterol side chains yields C-19 steroids, C-22 steroids, and 17-ketosteroids (i.e. precursors to adrenocortical hormones and contraceptives). The addition and modification of functional groups is key when producing the wide variety of medications available within this chemical classification. These modifications are performed using conventional organic synthesis and/or biotransformation techniques.
Semisynthesis
The semisynthesis of steroids often begins from precursors such as cholesterol, phytosterols, or sapogenins. The efforts of Syntex, a company involved in the Mexican barbasco trade, used Dioscorea mexicana to produce the sapogenin diosgenin in the early days of the synthetic steroid pharmaceutical industry.
Total synthesis
Some steroidal hormones are economically obtained only by total synthesis from petrochemicals (e.g. 13-alkyl steroids). For example, the pharmaceutical Norgestrel begins from Methoxy-1-tetralone, a petrochemical derived from phenol.
Research awards
A number of Nobel Prizes have been awarded for steroid research, including: