
Scientific name Vertebrata Higher classification Craniate | Clade Craniata Phylum Chordata Rank Subphylum | |
 | ||
Lower classifications Amphibians, Shark, Frog, Tetrapod, Chondrichthyes | ||
Vertebrate animals educational video for kids
Vertebrates /ˈvɜːrtᵻbrᵻts/ comprise all species of animals within the subphylum Vertebrata /-ɑː/ (chordates with backbones). Vertebrates represent the overwhelming majority of the phylum Chordata, with currently about 66,000 species described. Vertebrates include the jawless fish and the jawed vertebrates, which include the cartilaginous fish (sharks, rays, and ratfish) and the bony fish.
Contents
- Vertebrate animals educational video for kids
- Animal classification for children classifying vertebrates and invertebrates for kids freeschool
- Etymology
- Anatomy and morphology
- Vertebral column
- Gills
- Central nervous system
- First vertebrates
- From fish to amphibians
- Mesozoic vertebrates
- After the Mesozoic
- Classification
- Traditional classification
- Phylogenetic relationships
- Number of extant species
- Reproductive systems
- Inbreeding
- Inbreeding avoidance
- Outcrossing
- Parthenogenesis
- Self fertilization
- References
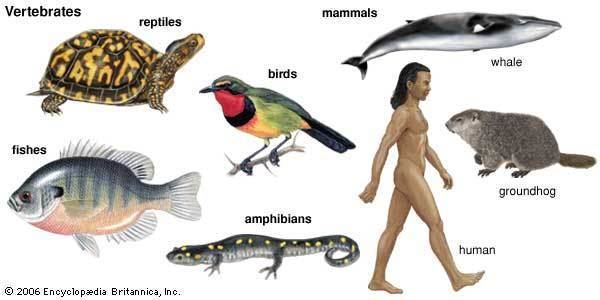
A bony fish clade known as the lobe-finned fishes is included with tetrapods, which are further divided into amphibians, reptiles, mammals, and birds. Extant vertebrates range in size from the frog species Paedophryne amauensis, at as little as 7.7 mm (0.30 in), to the blue whale, at up to 33 m (108 ft). Vertebrates make up less than five percent of all described animal species; the rest are invertebrates, which lack vertebral columns.

The vertebrates traditionally include the hagfish, which do not have proper vertebrae due to their loss in evolution, though their closest living relatives, the lampreys, do. Hagfish do, however, possess a cranium. For this reason, the vertebrate subphylum is sometimes referred to as "Craniata" when discussing morphology.
Molecular analysis since 1992 has suggested that hagfish are most closely related to lampreys, and so also are vertebrates in a monophyletic sense. Others consider them a sister group of vertebrates in the common taxon of craniata.
Animal classification for children classifying vertebrates and invertebrates for kids freeschool
Etymology
The word origin of vertebrate derives from the Latin word vertebratus (Pliny), meaning joint of the spine. The Proto-Indo-European language origins are still unclear.
Vertebrate is derived from the word vertebra, which refers to any of the bones or segments of the spinal column.
Anatomy and morphology
All vertebrates are built along the basic chordate body plan: a stiff rod running through the length of the animal (vertebral column and/or notochord), with a hollow tube of nervous tissue (the spinal cord) above it and the gastrointestinal tract below.
In all vertebrates, the mouth is found at, or right below, the anterior end of the animal, while the anus opens to the exterior before the end of the body. The remaining part of the body continuing after the anus forms a tail with vertebrae and spinal cord, but no gut.
Vertebral column
The defining characteristic of a vertebrate is the vertebral column, in which the notochord (a stiff rod of uniform composition) found in all chordates has been replaced by a segmented series of stiffer elements (vertebrae) separated by mobile joints (intervertebral discs, derived embryonically and evolutionarily from the notochord).
However, a few vertebrates have secondarily lost this anatomy, retaining the notochord into adulthood, such as the sturgeon and coelacanth. Jawed vertebrates are typified by paired appendages (fins or legs, which may be secondarily lost), but this trait is not required in order for an animal to be a vertebrate.
Gills
All basal vertebrates breathe with gills. The gills are carried right behind the head, bordering the posterior margins of a series of openings from the pharynx to the exterior. Each gill is supported by a cartilagenous or bony gill arch. The bony fish have three pairs of arches, cartilaginous fish have five to seven pairs, while the primitive jawless fish have seven. The vertebrate ancestor no doubt had more arches than this, as some of their chordate relatives have more than 50 pairs of gills.
In amphibians and some primitive bony fishes, the larvae bear external gills, branching off from the gill arches. These are reduced in adulthood, their function taken over by the gills proper in fishes and by lungs in most amphibians. Some amphibians retain the external larval gills in adulthood, the complex internal gill system as seen in fish apparently being irrevocably lost very early in the evolution of tetrapods.
While the higher vertebrates lack gills, the gill arches form during fetal development, and form the basis of essential structures such as jaws, the thyroid gland, the larynx, the columella (corresponding to the stapes in mammals) and, in mammals, the malleus and incus.
Central nervous system
The central nervous system of vertebrates is based on a hollow nerve cord running along the length of the animal. Of particular importance and unique to vertebrates is the presence of neural crest cells. These are progenitors of stem cells, and critical to coordinating the functions of cellular components. Neural crest cells migrate through the body from the nerve cord during development, and initiate the formation of neural ganglia and structures such as the jaws and skull.
The vertebrates are the only chordate group to exhibit cephalisation, the concentration of brain functions in the head. A slight swelling of the anterior end of the nerve cord is found in the lancelet, a chordate, though it lacks the eyes and other complex sense organs comparable to those of vertebrates. Other chordates do not show any trends towards cephalisation.
A peripheral nervous system branches out from the nerve cord to innervate the various systems. The front end of the nerve tube is expanded by a thickening of the walls and expansion of the central canal of spinal cord into three primary brain vesicles: The prosencephalon (forebrain), mesencephalon (midbrain) and rhombencephalon (hindbrain), further differentiated in the various vertebrate groups. Two laterally placed eyes form around outgrowths from the midbrain, except in hagfish, though this may be a secondary loss. The forebrain is well developed and subdivided in most tetrapods, while the midbrain dominate in many fish and some salamanders. Vesicles of the forebrain are usually paired, giving rise to hemispheres like the cerebral hemispheres in mammals.
The resulting anatomy of the central nervous system, with a single hollow nerve cord topped by a series of (often paired) vesicles, is unique to vertebrates. All invertebrates with well-developed brains, such as insects, spiders and squids, have a ventral rather than dorsal system of ganglions, with a split brain stem running on each side of the mouth or gut.
First vertebrates
Vertebrates originated about 525 million years ago during the Cambrian explosion, which saw the rise in organism diversity. The earliest known vertebrate is believed to be the Myllokunmingia. Another early vertebrate is Haikouichthys ercaicunensis. Unlike the other fauna that dominated the Cambrian, these groups had the basic vertebrate body plan: a notochord, rudimentary vertebrae, and a well-defined head and tail. All of these early vertebrates lacked jaws in the common sense and relied on filter feeding close to the seabed. A vertebrate group of uncertain phylogeny, small-eel-like conodonts, are known from microfossils of their paired tooth segments from the late Cambrian to the end of the Triassic.
From fish to amphibians
The first jawed vertebrates appeared in the latest Ordovician and became common in the Devonian, often known as the "Age of Fishes". The two groups of bony fishes, the actinopterygii and sarcopterygii, evolved and became common. The Devonian also saw the demise of virtually all jawless fishes, save for lampreys and hagfish, as well as the Placodermi, a group of armoured fish that dominated the entirety of that period since the late Silurian. The Devonian also saw the rise of the first labyrinthodonts, which was a transitional form between fishes and amphibians.
Mesozoic vertebrates
Amniotes branched from labyrinthodonts in the subsequent Carboniferous period. The Parareptilia and synapsid amniotes were common during the late Paleozoic, while diapsids became dominant during the Mesozoic. In the sea, the bony fishes became dominant. The birds are a derived form of dinosaurs evolved in the Jurassic. The demise of the non-avian dinosaurs at the end of the Cretaceous allowed for the expansion of mammals, which had evolved from the therapsids, a group of synapsid amniotes, during the late Triassic Period.
After the Mesozoic
The Cenozoic world has seen great diversification of bony fishes, frogs, birds and mammals.
Over half of all living vertebrate species (about 32,000 species) are fish (non-tetrapod craniates), a diverse set of lineages that inhabit all the world's aquatic ecosystems, from snow minnows (Cypriniformes) in Himalayan lakes at elevations over 4,600 metres (15,100 feet) to flatfishes (order Pleuronectiformes) in the Challenger Deep, the deepest ocean trench at about 11,000 metres (36,000 feet). Fishes of myriad varieties are the main predators in most of the world's water bodies, both freshwater and marine. The rest of the vertebrate species are tetrapods, a single lineage that includes amphibians (with roughly 7,000 species); mammals (with approximately 5,500 species); and reptiles and birds (with about 20,000 species divided evenly between the two classes). Tetrapods comprise the dominant megafauna of most terrestrial environments and also include many partially or fully aquatic groups (e.g., sea snakes, penguins, cetaceans).
Classification
There are several ways of classifying animals. Evolutionary systematics relies on anatomy, physiology and evolutionary history, which is determined through similarities in anatomy and, if possible, the genetics of organisms. Phylogenetic classification is based solely on phylogeny. Evolutionary systematics gives an overview; phylogenetic systematics gives detail. The two systems are thus complementary rather than opposed.
Traditional classification
Conventional classification has living vertebrates grouped into seven classes based on traditional interpretations of gross anatomical and physiological traits. This classification is the one most commonly encountered in school textbooks, overviews, non-specialist, and popular works. The extant vertebrates are:
In addition to these, there are two classes of extinct armoured fishes, the Placodermi and the Acanthodii.
Other ways of classifying the vertebrates have been devised, particularly with emphasis on the phylogeny of early amphibians and reptiles. An example based on Janvier (1981, 1997), Shu et al. (2003), and Benton (2004) is given here:
While this traditional classification is orderly, most of the groups are paraphyletic, i.e. do not contain all descendants of the class's common ancestor. For instance, descendants of the first reptiles include modern reptiles, as well as mammals and birds. Most of the classes listed are not "complete" (and are therefore paraphyletic) taxa, meaning they do not include all the descendants of the first representative of the group. For example, the agnathans have given rise to the jawed vertebrates; the bony fishes have given rise to the land vertebrates; the traditional "amphibians" have given rise to the reptiles (traditionally including the synapsids, or mammal-like "reptiles"), which in turn have given rise to the mammals and birds. Most scientists working with vertebrates use a classification based purely on phylogeny, organized by their known evolutionary history and sometimes disregarding the conventional interpretations of their anatomy and physiology.
Phylogenetic relationships
In phylogenetic taxonomy, the relationships between animals are not typically divided into ranks, but illustrated as a nested "family tree" known as a cladogram. Phylogenetic groups are given definitions based on their relationship to one another, rather than purely on physical traits, such as the presence of a backbone. This nesting pattern is often combined with traditional taxonomy (as above), in a practice known as evolutionary taxonomy.
The cladogram presented below is based on studies compiled by Philippe Janvier and others for the Tree of Life Web Project.
Number of extant species
The number of described vertebrate species are split evenly between tetrapods and fish. The following table lists the number of described extant species for each vertebrate class as estimated in the IUCN Red List of Threatened Species, 2014.3.
The IUCN estimates that 1,305,075 extant invertebrate species have been described, which means that less than 5% of the described animal species in the world are vertebrates.
Reproductive systems
Nearly all vertebrates undergo sexual reproduction. They produce haploid gametes by meiosis. The smaller, motile gametes are spermatozoa and the larger, non-motile gametes are ova. These fuse by the process of fertilisation to form diploid zygotes, which develop into new individuals.
Inbreeding
During sexual reproduction, mating with a close relative (inbreeding) often leads to inbreeding depression. Inbreeding depression is considered to be largely due to expression of deleterious recessive mutations. The effects of inbreeding have been studied in many vertebrate species.
In several species of fish, inbreeding was found to decrease reproductive success.
Inbreeding was observed to increase juvenile mortality in 11 small animal species.
A common breeding practice for pet dogs is mating between close relatives (e.g. between half- and full siblings). This practice generally has a negative effect on measures of reproductive success, including decreased litter size and puppy survival.
Incestuous matings in birds result in severe fitness costs due to inbreeding depression (e.g. reduction in hatchability of eggs and reduced progeny survival).
Inbreeding avoidance
As a result of the negative fitness consequences of inbreeding, vertebrate species have evolved mechanisms to avoid inbreeding. Numerous inbreeding avoidance mechanisms operating prior to mating have been described.
Toads and many other amphibians display breeding site fidelity. Individuals that return to natal ponds to breed will likely encounter siblings as potential mates. Although incest is possible, Bufo americanus siblings rarely mate. These toads likely recognize and actively avoid close kins as mates. Advertisement vocalizations by males appear to serve as cues by which females recognize their kin.
Inbreeding avoidance mechanisms can also operate subsequent to copulation. In guppies, a post-copulatory mechanism of inbreeding avoidance occurs based on competition between sperm of rival males for achieving fertilization. In competitions between sperm from an unrelated male and from a full sibling male, a significant bias in paternity towards the unrelated male was observed.
When female sand lizards mate with two or more males, sperm competition within the female's reproductive tract may occur. Active selection of sperm by females appears to occur in a manner that enhances female fitness. On the basis of this selective process, the sperm of males that are more distantly related to the female are preferentially used for fertilization, rather than the sperm of close relatives. This preference may enhance the fitness of progeny by reducing inbreeding depression.
Outcrossing
Mating with unrelated or distantly related members of the same species is generally thought to provide the advantage of masking deleterious recessive mutations in progeny (and see Heterosis). Vertebrates have evolved numerous diverse mechanisms for avoiding close inbreeding and promoting outcrossing (and see Inbreeding avoidance).
Outcrossing as a way of avoiding inbreeding depression, has been especially well studied in birds. For instance, inbreeding depression occurs in the great tit when the offspring are produced as a result of a mating between close relatives. In natural populations of the great tit (Parus major), inbreeding is avoided by dispersal of individuals from their birthplace, which reduces the chance of mating with a close relative.
The purple-crowned fairywren females paired with related males may undertake extra-pair matings that can reduce the negative effects of inbreeding. However, there are ecological and demographic constraints on extra pair matings. Nevertheless, 46% of broods produced by incestuously paired females contained extra-pair young.
Southern pied babblers (Turdoides bicolor) appear to avoid inbreeding in two ways. The first is through dispersal, and the second is by avoiding familiar group members as mates. Although both males and females disperse locally, they move outside the range where genetically related individuals are likely to be encountered. Within their group, individuals only acquire breeding positions when the opposite-sex breeder is unrelated.
Cooperative breeding in birds typically occurs when offspring, usually males, delay dispersal from their natal group in order to remain with the family to help rear younger kin. Female offspring rarely stay at home, dispersing over distances that allow them to breed independently, or to join unrelated groups.
Parthenogenesis
Parthenogenesis is a natural form of reproduction in which growth and development of embryos occur without fertilization.
Reproduction in squamate reptiles is ordinarily sexual, with males having a ZZ pair of sex determining chromosomes, and females a ZW pair. However, various species, including the Colombian Rainbow boa (Epicrates maurus), Agkistrodon contortrix (copperhead snake) and Agkistrodon piscivorus (cotton mouth snake) can also reproduce by facultative parthenogenesis -that is, they are capable of switching from a sexual mode of reproduction to an asexual mode- resulting in production of WW female progeny. The WW females are likely produced by terminal automixis.
Mole salamanders are an ancient (2.4-3.8 million year-old) unisexual vertebrate lineage. In the polyploid unisexual mole salamander females, a premeiotic endomitotic event doubles the number of chromosomes. As a result, the mature eggs produced subsequent to the two meiotic divisions have the same ploidy as the somatic cells of the female salamander. Synapsis and recombination during meiotic prophase I in these unisexual females is thought to ordinarily occur between identical sister chromosomes and occasionally between homologous chromosomes. Thus little, if any, genetic variation is produced. Recombination between homeologous chromosomes occurs only rarely, if at all. Since production of genetic variation is weak, at best, it is unlikely to provide a benefit sufficient to account for the long-term maintenance of meiosis in these organisms. However, meiosis may have been maintained during evolution by the efficient recombinational repair of DNA damages that meiosis provides, an advantage that could be realized at each generation.
Self-fertilization
The mangrove killifish (Kryptolebias marmoratus) produces both eggs and sperm by meiosis and routinely reproduces by self-fertilisation. The capacity for selfing in these fishes has apparently persisted for at least several hundred thousand years. Each individual hermaphrodite normally fertilizes itself when an egg and sperm that it has produced by an internal organ unite inside the fish's body. In nature, this mode of reproduction can yield highly homozygous lines composed of individuals so genetically uniform as to be, in effect, identical to one another. Although inbreeding, especially in the extreme form of self-fertilization, is ordinarily regarded as detrimental because it leads to expression of deleterious recessive alleles, self-fertilization does provide the benefit of fertilization assurance (reproductive assurance) at each generation.