
 | ||
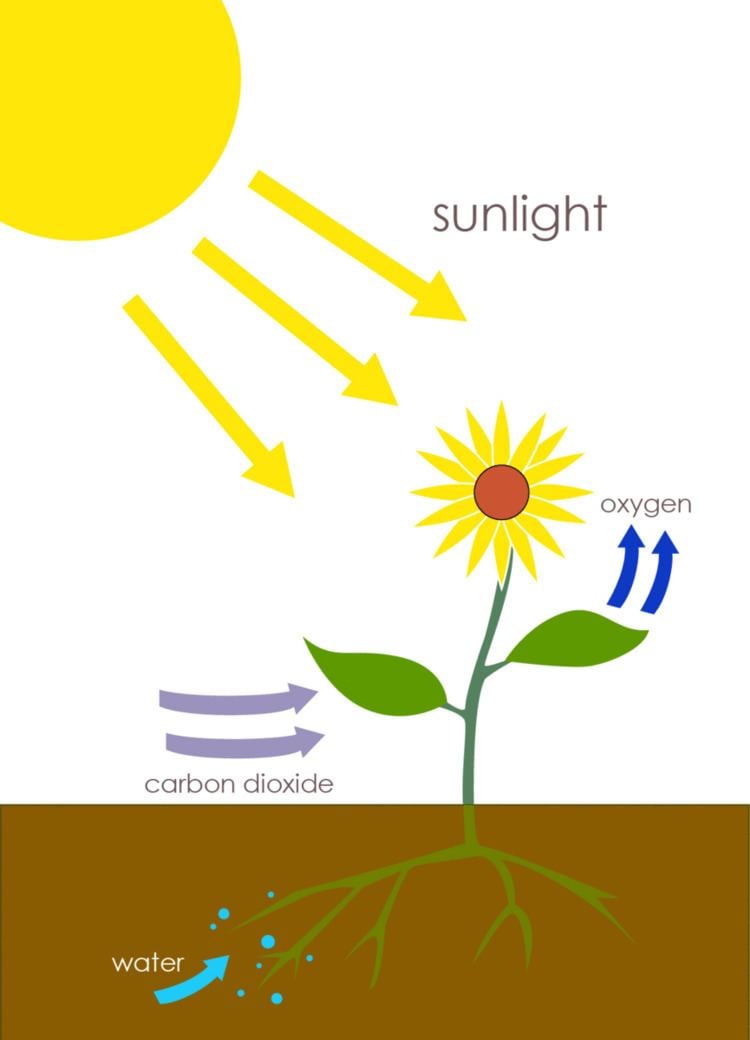
Photosynthesis is a process used by plants and other organisms to convert light energy into chemical energy that can later be released to fuel the organisms' activities (energy transformation). This chemical energy is stored in carbohydrate molecules, such as sugars, which are synthesized from carbon dioxide and water – hence the name photosynthesis, from the Greek φῶς, phōs, "light", and σύνθεσις, synthesis, "putting together". In most cases, oxygen is also released as a waste product. Most plants, most algae, and cyanobacteria perform photosynthesis; such organisms are called photoautotrophs. Photosynthesis is largely responsible for producing and maintaining the oxygen content of the Earth's atmosphere, and supplies all of the organic compounds and most of the energy necessary for life on Earth.
Contents
- Overview
- Photosynthetic membranes and organelles
- Light dependent reactions
- Z scheme
- Water photolysis
- Calvin cycle
- On land
- In water
- Order and kinetics
- Efficiency
- Evolution
- Symbiosis and the origin of chloroplasts
- Cyanobacteria and the evolution of photosynthesis
- Discovery
- Development of the concept
- C3 C4 photosynthesis research
- Factors
- Light intensity irradiance wavelength and temperature
- Carbon dioxide levels and photorespiration
- References
Although photosynthesis is performed differently by different species, the process always begins when energy from light is absorbed by proteins called reaction centres that contain green chlorophyll pigments. In plants, these proteins are held inside organelles called chloroplasts, which are most abundant in leaf cells, while in bacteria they are embedded in the plasma membrane. In these light-dependent reactions, some energy is used to strip electrons from suitable substances, such as water, producing oxygen gas. The hydrogen freed by the splitting of water is used in the creation of two further compounds that act as an immediate energy storage means: reduced nicotinamide adenine dinucleotide phosphate (NADPH) and adenosine triphosphate (ATP), the "energy currency" of cells.
In plants, algae and cyanobacteria, long-term energy storage in the form of sugars is produced by a subsequent sequence of light-independent reactions called the Calvin cycle; some bacteria use different mechanisms, such as the reverse Krebs cycle, to achieve the same end. In the Calvin cycle, atmospheric carbon dioxide is incorporated into already existing organic carbon compounds, such as ribulose bisphosphate (RuBP). Using the ATP and NADPH produced by the light-dependent reactions, the resulting compounds are then reduced and removed to form further carbohydrates, such as glucose.
The first photosynthetic organisms probably evolved early in the evolutionary history of life and most likely used reducing agents such as hydrogen or hydrogen sulfide, rather than water, as sources of electrons. Cyanobacteria appeared later; the excess oxygen they produced contributed directly to the oxygenation of the Earth, which rendered the evolution of complex life possible. Today, the average rate of energy capture by photosynthesis globally is approximately 130 terawatts, which is about three times the current power consumption of human civilization. Photosynthetic organisms also convert around 100–115 thousand million metric tonnes of carbon into biomass per year.
Overview
Photosynthetic organisms are photoautotrophs, which means that they are able to synthesize food directly from carbon dioxide and water using energy from light. However, not all organisms that use light as a source of energy carry out photosynthesis; photoheterotrophs use organic compounds, rather than carbon dioxide, as a source of carbon. In plants, algae, and cyanobacteria, photosynthesis releases oxygen. This is called oxygenic photosynthesis and is by far the most common type of photosynthesis used by living organisms. Although there are some differences between oxygenic photosynthesis in plants, algae, and cyanobacteria, the overall process is quite similar in these organisms. There are also many varieties of anoxygenic photosynthesis, used mostly by certain types of bacteria, which consume carbon dioxide but do not release oxygen.
Carbon dioxide is converted into sugars in a process called carbon fixation. Photosynthesis provides the energy in the form of free electrons that are used to split carbon from carbon dioxide that is then used to fix that carbon once again as carbohydrate. Carbon fixation is an endothermic redox reaction, so photosynthesis supplies the energy that drives both process. In general outline, photosynthesis is the opposite of cellular respiration, in which glucose and other compounds are oxidized to produce carbon dioxide and water, and to release chemical energy (an exothermic reaction) to drive the organism's metabolism. The two processes, reduction of carbon dioxide to carbohydrate and then later oxidation of the carbohydrate, are distinct: photosynthesis and cellular respiration take place through a different sequence of chemical reactions and in different cellular compartments.
The general equation for photosynthesis as first proposed by Cornelius van Niel is therefore:
CO2 + 2H2A + photons → [CH2O] + 2A + H2Ocarbon dioxide + electron donor + light energy → carbohydrate + oxidized electron donor + waterSince water is used as the electron donor in oxygenic photosynthesis, the equation for this process is:
CO2 + 2H2O + photons → [CH2O] + O2 + H2Ocarbon dioxide + water + light energy → carbohydrate + oxygen + waterThis equation emphasizes that water is both a reactant in the light-dependent reaction and a product of the light-independent reaction, but canceling n water molecules from each side gives the net equation:
CO2 + H2O + photons → [CH2O] + O2carbon dioxide + water + light energy → carbohydrate + oxygenOther processes substitute other compounds (such as arsenite) for water in the electron-supply role; for example some microbes use sunlight to oxidize arsenite to arsenate: The equation for this reaction is:
CO2 + (AsO3−3) + photons → (AsO3−
4) + COcarbon dioxide + arsenite + light energy → arsenate + carbon monoxide (used to build other compounds in subsequent reactions)
Photosynthesis occurs in two stages. In the first stage, light-dependent reactions or light reactions capture the energy of light and use it to make the energy-storage molecules ATP and NADPH. During the second stage, the light-independent reactions use these products to capture and reduce carbon dioxide.
Most organisms that utilize oxygenic photosynthesis use visible light for the light-dependent reactions, although at least three use shortwave infrared or, more specifically, far-red radiation.
Some organisms employ even more radical variants of photosynthesis. Some archea use a simpler method that employs a pigment similar to those used for vision in animals. The bacteriorhodopsin changes its configuration in response to sunlight, acting as a proton pump. This produces a proton gradient more directly, which is then converted to chemical energy. The process does not involve carbon dioxide fixation and does not release oxygen, and seems to have evolved separately from the more common types of photosynthesis.
Photosynthetic membranes and organelles
In photosynthetic bacteria, the proteins that gather light for photosynthesis are embedded in cell membranes. In its simplest form, this involves the membrane surrounding the cell itself. However, the membrane may be tightly folded into cylindrical sheets called thylakoids, or bunched up into round vesicles called intracytoplasmic membranes. These structures can fill most of the interior of a cell, giving the membrane a very large surface area and therefore increasing the amount of light that the bacteria can absorb.
In plants and algae, photosynthesis takes place in organelles called chloroplasts. A typical plant cell contains about 10 to 100 chloroplasts. The chloroplast is enclosed by a membrane. This membrane is composed of a phospholipid inner membrane, a phospholipid outer membrane, and an intermembrane space. Enclosed by the membrane is an aqueous fluid called the stroma. Embedded within the stroma are stacks of thylakoids (grana), which are the site of photosynthesis. The thylakoids appear as flattened disks. The thylakoid itself is enclosed by the thylakoid membrane, and within the enclosed volume is a lumen or thylakoid space. Embedded in the thylakoid membrane are integral and peripheral membrane protein complexes of the photosynthetic system.
Plants absorb light primarily using the pigment chlorophyll. The green part of the light spectrum is not absorbed but is reflected which is the reason that most plants have a green color. Besides chlorophyll, plants also use pigments such as carotenes and xanthophylls. Algae also use chlorophyll, but various other pigments are present, such as phycocyanin, carotenes, and xanthophylls in green algae, phycoerythrin in red algae (rhodophytes) and fucoxanthin in brown algae and diatoms resulting in a wide variety of colors.
These pigments are embedded in plants and algae in complexes called antenna proteins. In such proteins, the pigments are arranged to work together. Such a combination of proteins is also called a light-harvesting complex.
Although all cells in the green parts of a plant have chloroplasts, the majority of those are found in specially adapted structures called leaves. Certain species adapted to conditions of strong sunlight and aridity, such as many Euphorbia and cactus species, have their main photosynthetic organs in their stems. The cells in the interior tissues of a leaf, called the mesophyll, can contain between 450,000 and 800,000 chloroplasts for every square millimeter of leaf. The surface of the leaf is coated with a water-resistant waxy cuticle that protects the leaf from excessive evaporation of water and decreases the absorption of ultraviolet or blue light to reduce heating. The transparent epidermis layer allows light to pass through to the palisade mesophyll cells where most of the photosynthesis takes place.
Light-dependent reactions
In the light-dependent reactions, one molecule of the pigment chlorophyll absorbs one photon and loses one electron. This electron is passed to a modified form of chlorophyll called pheophytin, which passes the electron to a quinone molecule, starting the flow of electrons down an electron transport chain that leads to the ultimate reduction of NADP to NADPH. In addition, this creates a proton gradient (energy gradient) across the chloroplast membrane, which is used by ATP synthase in the synthesis of ATP. The chlorophyll molecule ultimately regains the electron it lost when a water molecule is split in a process called photolysis, which releases a dioxygen (O2) molecule as a waste product.
The overall equation for the light-dependent reactions under the conditions of non-cyclic electron flow in green plants is:
2 H2O + 2 NADP+ + 3 ADP + 3 Pi + light → 2 NADPH + 2 H+ + 3 ATP + O2Not all wavelengths of light can support photosynthesis. The photosynthetic action spectrum depends on the type of accessory pigments present. For example, in green plants, the action spectrum resembles the absorption spectrum for chlorophylls and carotenoids with absorption peaks in violet-blue and red light. In red algae, the action spectrum is blue-green light, which allows these algae to use the blue end of the spectrum to grow in the deeper waters that filter out the longer wavelengths (red light) used by above ground green plants. The non-absorbed part of the light spectrum is what gives photosynthetic organisms their color (e.g., green plants, red algae, purple bacteria) and is the least effective for photosynthesis in the respective organisms.
Z scheme
In plants, light-dependent reactions occur in the thylakoid membranes of the chloroplasts where they drive the synthesis of ATP and NADPH. The light-dependent reactions are of two forms: cyclic and non-cyclic.
In the non-cyclic reaction, the photons are captured in the light-harvesting antenna complexes of photosystem II by chlorophyll and other accessory pigments (see diagram at right). The absorption of a photon by the antenna complex frees an electron by a process called photoinduced charge separation. The antenna system is at the core of the chlorophyll molecule of the photosystem II reaction center. That freed electron is transferred to the primary electron-acceptor molecule, pheophytin. As the electrons are shuttled through an electron transport chain (the so-called Z-scheme shown in the diagram), it initially functions to generate a chemiosmotic potential by pumping proton cations (H+) across the membrane and into the thylakoid space. An ATP synthase enzyme uses that chemiosmotic potential to make ATP during photophosphorylation, whereas NADPH is a product of the terminal redox reaction in the Z-scheme. The electron enters a chlorophyll molecule in Photosystem I. There it is further excited by the light absorbed by that photosystem. The electron is then passed along a chain of electron acceptors to which it transfers some of its energy. The energy delivered to the electron acceptors is used to move hydrogen ions across the thylakoid membrane into the lumen. The electron is eventually used to reduce the co-enzyme NADP with a H+ to NADPH (which has functions in the light-independent reaction); at that point, the path of that electron ends.
The cyclic reaction is similar to that of the non-cyclic, but differs in that it generates only ATP, and no reduced NADP (NADPH) is created. The cyclic reaction takes place only at photosystem I. Once the electron is displaced from the photosystem, the electron is passed down the electron acceptor molecules and returns to photosystem I, from where it was emitted, hence the name cyclic reaction.
Water photolysis
The NADPH is the main reducing agent produced by chloroplasts, which then goes on to provide a source of energetic electrons in other cellular reactions. Its production leaves chlorophyll in photosystem I with a deficit of electrons (chlorophyll has been oxidized), which must be balanced by some other reducing agent that will supply the missing electron. The excited electrons lost from chlorophyll from photosystem I are supplied from the electron transport chain by plastocyanin. However, since photosystem II is the first step of the Z-scheme, an external source of electrons is required to reduce its oxidized chlorophyll a molecules. The source of electrons in green-plant and cyanobacterial photosynthesis is water. Two water molecules are oxidized by four successive charge-separation reactions by photosystem II to yield a molecule of diatomic oxygen and four hydrogen ions; the electrons yielded are transferred to a redox-active tyrosine residue that then reduces the oxidized chlorophyll a (called P680+) that serves as the primary light-driven electron donor in the photosystem II reaction center. That photo receptor is in effect reset and is then able to repeat the absorption of another photon and the release of another photo-dissociated electron. The oxidation of water is catalyzed in photosystem II by a redox-active structure that contains four manganese ions and a calcium ion; this oxygen-evolving complex binds two water molecules and contains the four oxidizing equivalents that are used to drive the water-oxidizing reaction. Photosystem II is the only known biological enzyme that carries out this oxidation of water. The hydrogen ions released contribute to the transmembrane chemiosmotic potential that leads to ATP synthesis. Oxygen is a waste product of light-dependent reactions, but the majority of organisms on Earth use oxygen for cellular respiration, including photosynthetic organisms.
Calvin cycle
In the light-independent (or "dark") reactions, the enzyme RuBisCO captures CO2 from the atmosphere and, in a process called the Calvin-Benson cycle, it uses the newly formed NADPH and releases three-carbon sugars, which are later combined to form sucrose and starch. The overall equation for the light-independent reactions in green plants is
3 CO2 + 9 ATP + 6 NADPH + 6 H+ → C3H6O3-phosphate + 9 ADP + 8 Pi + 6 NADP+ + 3 H2OCarbon fixation produces the intermediate three-carbon sugar product, which is then converted to the final carbohydrate products. The simple carbon sugars produced by photosynthesis are then used in the forming of other organic compounds, such as the building material cellulose, the precursors for lipid and amino acid biosynthesis, or as a fuel in cellular respiration. The latter occurs not only in plants but also in animals when the energy from plants is passed through a food chain.
The fixation or reduction of carbon dioxide is a process in which carbon dioxide combines with a five-carbon sugar, ribulose 1,5-bisphosphate, to yield two molecules of a three-carbon compound, glycerate 3-phosphate, also known as 3-phosphoglycerate. Glycerate 3-phosphate, in the presence of ATP and NADPH produced during the light-dependent stages, is reduced to glyceraldehyde 3-phosphate. This product is also referred to as 3-phosphoglyceraldehyde (PGAL) or, more generically, as triose phosphate. Most (5 out of 6 molecules) of the glyceraldehyde 3-phosphate produced is used to regenerate ribulose 1,5-bisphosphate so the process can continue. The triose phosphates not thus "recycled" often condense to form hexose phosphates, which ultimately yield sucrose, starch and cellulose. The sugars produced during carbon metabolism yield carbon skeletons that can be used for other metabolic reactions like the production of amino acids and lipids.
On land
In hot and dry conditions, plants close their stomata to prevent water loss. Under these conditions, CO2 will decrease and oxygen gas, produced by the light reactions of photosynthesis, will increase, causing an increase of photorespiration by the oxygenase activity of ribulose-1,5-bisphosphate carboxylase/oxygenase and decrease in carbon fixation. Some plants have evolved mechanisms to increase the CO2 concentration in the leaves under these conditions.
Plants that use the C4 carbon fixation process chemically fix carbon dioxide in the cells of the mesophyll by adding it to the three-carbon molecule phosphoenolpyruvate (PEP), a reaction catalyzed by an enzyme called PEP carboxylase, creating the four-carbon organic acid oxaloacetic acid. Oxaloacetic acid or malate synthesized by this process is then translocated to specialized bundle sheath cells where the enzyme RuBisCO and other Calvin cycle enzymes are located, and where CO2 released by decarboxylation of the four-carbon acids is then fixed by RuBisCO activity to the three-carbon 3-phosphoglyceric acids. The physical separation of RuBisCO from the oxygen-generating light reactions reduces photorespiration and increases CO2 fixation and, thus, the photosynthetic capacity of the leaf. C4 plants can produce more sugar than C3 plants in conditions of high light and temperature. Many important crop plants are C4 plants, including maize, sorghum, sugarcane, and millet. Plants that do not use PEP-carboxylase in carbon fixation are called C3 plants because the primary carboxylation reaction, catalyzed by RuBisCO, produces the three-carbon 3-phosphoglyceric acids directly in the Calvin-Benson cycle. Over 90% of plants use C3 carbon fixation, compared to 3% that use C4 carbon fixation; however, the evolution of C4 in over 60 plant lineages makes it a striking example of convergent evolution.
Xerophytes, such as cacti and most succulents, also use PEP carboxylase to capture carbon dioxide in a process called Crassulacean acid metabolism (CAM). In contrast to C4 metabolism, which spatially separates the CO2 fixation to PEP from the Calvin cycle, CAM temporally separates these two processes. CAM plants have a different leaf anatomy from C3 plants, and fix the CO2 at night, when their stomata are open. CAM plants store the CO2 mostly in the form of malic acid via carboxylation of phosphoenolpyruvate to oxaloacetate, which is then reduced to malate. Decarboxylation of malate during the day releases CO2 inside the leaves, thus allowing carbon fixation to 3-phosphoglycerate by RuBisCO. Sixteen thousand species of plants use CAM.
In water
Cyanobacteria possess carboxysomes, which increase the concentration of CO2 around RuBisCO to increase the rate of photosynthesis. An enzyme, carbonic anhydrase, located within the carboxysome releases CO2 from the dissolved hydrocarbonate ions (HCO−
3). Before the CO2 diffuses out it is quickly sponged up by RuBisCO, which is concentrated within the carboxysomes. HCO−
3 ions are made from CO2 outside the cell by another carbonic anhydrase and are actively pumped into the cell by a membrane protein. They cannot cross the membrane as they are charged, and within the cytosol they turn back into CO2 very slowly without the help of carbonic anhydrase. This causes the HCO−
3 ions to accumulate within the cell from where they diffuse into the carboxysomes. Pyrenoids in algae and hornworts also act to concentrate CO2 around rubisco.
Order and kinetics
The overall process of photosynthesis takes place in four stages:
Efficiency
Plants usually convert light into chemical energy with a photosynthetic efficiency of 3–6%. Absorbed light that is unconverted is dissipated primarily as heat, with a small fraction (1–2%) re-emitted as chlorophyll fluorescence at longer (redder) wavelengths. A fact that allows measurement of the light reaction of photosynthesis by using chlorophyll fluorometers.
Actual plants' photosynthetic efficiency varies with the frequency of the light being converted, light intensity, temperature and proportion of carbon dioxide in the atmosphere, and can vary from 0.1% to 8%. By comparison, solar panels convert light into electric energy at an efficiency of approximately 6–20% for mass-produced panels, and above 40% in laboratory devices.
The efficiency of both light and dark reactions can be measured but the relationship between the two can be complex. For example, the ATP and NADPH energy molecules, created by the light reaction, can be used for carbon fixation or for photorespiration in C3 plants. Electrons may also flow to other electron sinks. For this reason, it is not uncommon for authors to differentiate between work done under non-photorespiratory conditions and under photorespiratory conditions.
Chlorophyll fluorescence of photosystem II can measure the light reaction, and Infrared gas analyzers can measure the dark reaction. It is also possible to investigate both at the same time using an integrated chlorophyll fluorometer and gas exchange system, or by using two separate systems together. Infrared gas analyzers and some moisture sensors are sensitive enough to measure the photosynthetic assimilation of CO2, and of ΔH2O using reliable methods CO2 is commonly measured in μmols/m2/s−1, parts per million or volume per million and H20 is commonly measured in mmol/m2/s−1 or in mbars. By measuring CO2 assimilation, ΔH2O, leaf temperature, barometric pressure, leaf area, and photosynthetically active radiation or PAR, it becomes possible to estimate, “A” or carbon assimilation, “E” or transpiration, “gs” or stomatal conductance, and Ci or intracellular CO2. However, it is more common to used chlorophyll fluorescence for plant stress measurement, where appropriate, because the most commonly used measuring parameters FV/FM and Y(II) or F/FM’ can be made in a few seconds, allowing the measurement of larger plant populations.
Gas exchange systems that offer control of CO2 levels, above and below ambient, allow the common practice of measurement of A/Ci curves, at different CO2 levels, to characterize a plant’s photosynthetic response.
Integrated chlorophyll fluorometer – gas exchange systems allow a more precise measure of photosynthetic response and mechanisms. While standard gas exchange photosynthesis systems can measure Ci, or substomatal CO2 levels, the addition of integrated chlorophyll fluorescence measurements allows a more precise measurement of CC to replace Ci. The estimation of CO2 at the site of carboxylation in the chloroplast, or CC, becomes possible with the measurement of mesophyll conductance or gm using an integrated system.
Photosynthesis measurement systems are not designed to directly measure the amount of light absorbed by the leaf. But analysis of chlorophyll-fluorescence, P700- and P515-absorbance and gas exchange measurements reveal detailed information about e.g. the photosystems, quantum efficiency and the CO2 assimilation rates. With some instruments even wavelength-dependency of the photosynthetic efficiency can be analyzed.
A phenomenon known as quantum walk increases the efficiency of the energy transport of light significantly. In the photosynthetic cell of an algae, bacterium, or plant, there are light-sensitive molecules called chromophores arranged in an antenna-shaped structure named a photocomplex. When a photon is absorbed by a chromophore, it is converted into a quasiparticle referred to as an exciton, which jumps from chromophore to chromophore towards the reaction center of the photocomplex, a collection of molecules that traps its energy in a chemical form that makes it accessible for the cell's metabolism. The exciton's wave properties enable it to cover a wider area and try out several possible paths simultaneously, allowing it to instantaneously "choose" the most efficient route, where it will have the highest probability of arriving at its destination in the minimum possible time. Because that quantum walking takes place at temperatures far higher than quantum phenomena usually occur, it is only possible over very short distances, due to obstacles in the form of destructive interference that come into play. These obstacles cause the particle to lose its wave properties for an instant before it regains them once again after it is freed from its locked position through a classic "hop". The movement of the electron towards the photo center is therefore covered in a series of conventional hops and quantum walks.
Evolution
Early photosynthetic systems, such as those in green and purple sulfur and green and purple nonsulfur bacteria, are thought to have been anoxygenic, and used various other molecules as electron donors rather than water. Green and purple sulfur bacteria are thought to have used hydrogen and sulfur as electron donors. Green nonsulfur bacteria used various amino and other organic acids as an electron donor. Purple nonsulfur bacteria used a variety of nonspecific organic molecules. The use of these molecules is consistent with the geological evidence that Earth's early atmosphere was highly reducing at that time.
Fossils of what are thought to be filamentous photosynthetic organisms have been dated at 3.4 billion years old.
The main source of oxygen in the Earth's atmosphere derives from oxygenic photosynthesis, and its first appearance is sometimes referred to as the oxygen catastrophe. Geological evidence suggests that oxygenic photosynthesis, such as that in cyanobacteria, became important during the Paleoproterozoic era around 2 billion years ago. Modern photosynthesis in plants and most photosynthetic prokaryotes is oxygenic. Oxygenic photosynthesis uses water as an electron donor, which is oxidized to molecular oxygen (O
2) in the photosynthetic reaction center.
Symbiosis and the origin of chloroplasts
Several groups of animals have formed symbiotic relationships with photosynthetic algae. These are most common in corals, sponges and sea anemones. It is presumed that this is due to the particularly simple body plans and large surface areas of these animals compared to their volumes. In addition, a few marine mollusks Elysia viridis and Elysia chlorotica also maintain a symbiotic relationship with chloroplasts they capture from the algae in their diet and then store in their bodies. This allows the mollusks to survive solely by photosynthesis for several months at a time. Some of the genes from the plant cell nucleus have even been transferred to the slugs, so that the chloroplasts can be supplied with proteins that they need to survive.
An even closer form of symbiosis may explain the origin of chloroplasts. Chloroplasts have many similarities with photosynthetic bacteria, including a circular chromosome, prokaryotic-type ribosome, and similar proteins in the photosynthetic reaction center. The endosymbiotic theory suggests that photosynthetic bacteria were acquired (by endocytosis) by early eukaryotic cells to form the first plant cells. Therefore, chloroplasts may be photosynthetic bacteria that adapted to life inside plant cells. Like mitochondria, chloroplasts possess their own DNA, separate from the nuclear DNA of their plant host cells and the genes in this chloroplast DNA resemble those found in cyanobacteria. DNA in chloroplasts codes for redox proteins such as those found in the photosynthetic reaction centers. The CoRR Hypothesis proposes that this Co-location is required for Redox Regulation.
Cyanobacteria and the evolution of photosynthesis
The biochemical capacity to use water as the source for electrons in photosynthesis evolved once, in a common ancestor of extant cyanobacteria. The geological record indicates that this transforming event took place early in Earth's history, at least 2450–2320 million years ago (Ma), and, it is speculated, much earlier. Because the Earth's atmosphere contained almost no oxygen during the estimated development of photosynthesis, it is believed that the first photosynthetic cyanobacteria did not generate oxygen. Available evidence from geobiological studies of Archean (>2500 Ma) sedimentary rocks indicates that life existed 3500 Ma, but the question of when oxygenic photosynthesis evolved is still unanswered. A clear paleontological window on cyanobacterial evolution opened about 2000 Ma, revealing an already-diverse biota of blue-green algae. Cyanobacteria remained the principal primary producers of oxygen throughout the Proterozoic Eon (2500–543 Ma), in part because the redox structure of the oceans favored photoautotrophs capable of nitrogen fixation. Green algae joined blue-green algae as the major primary producers of oxygen on continental shelves near the end of the Proterozoic, but it was only with the Mesozoic (251–65 Ma) radiations of dinoflagellates, coccolithophorids, and diatoms did the primary production of oxygen in marine shelf waters take modern form. Cyanobacteria remain critical to marine ecosystems as primary producers of oxygen in oceanic gyres, as agents of biological nitrogen fixation, and, in modified form, as the plastids of marine algae.
Discovery
Although some of the steps in photosynthesis are still not completely understood, the overall photosynthetic equation has been known since the 19th century.
Jan van Helmont began the research of the process in the mid-17th century when he carefully measured the mass of the soil used by a plant and the mass of the plant as it grew. After noticing that the soil mass changed very little, he hypothesized that the mass of the growing plant must come from the water, the only substance he added to the potted plant. His hypothesis was partially accurate — much of the gained mass also comes from carbon dioxide as well as water. However, this was a signaling point to the idea that the bulk of a plant's biomass comes from the inputs of photosynthesis, not the soil itself.
Joseph Priestley, a chemist and minister, discovered that, when he isolated a volume of air under an inverted jar, and burned a candle in it, the candle would burn out very quickly, much before it ran out of wax. He further discovered that a mouse could similarly "injure" air. He then showed that the air that had been "injured" by the candle and the mouse could be restored by a plant.
In 1778, Jan Ingenhousz, repeated Priestley's experiments. He discovered that it was the influence of sunlight on the plant that could cause it to revive a mouse in a matter of hours.
In 1796, Jean Senebier, a Swiss pastor, botanist, and naturalist, demonstrated that green plants consume carbon dioxide and release oxygen under the influence of light. Soon afterward, Nicolas-Théodore de Saussure showed that the increase in mass of the plant as it grows could not be due only to uptake of CO2 but also to the incorporation of water. Thus, the basic reaction by which photosynthesis is used to produce food (such as glucose) was outlined.
Cornelis Van Niel made key discoveries explaining the chemistry of photosynthesis. By studying purple sulfur bacteria and green bacteria he was the first to demonstrate that photosynthesis is a light-dependent redox reaction, in which hydrogen reduces carbon dioxide.
Robert Emerson discovered two light reactions by testing plant productivity using different wavelengths of light. With the red alone, the light reactions were suppressed. When blue and red were combined, the output was much more substantial. Thus, there were two photosystems, one absorbing up to 600 nm wavelengths, the other up to 700 nm. The former is known as PSII, the latter is PSI. PSI contains only chlorophyll "a", PSII contains primarily chlorophyll "a" with most of the available chlorophyll "b", among other pigment. These include phycobilins, which are the red and blue pigments of red and blue algae respectively, and fucoxanthol for brown algae and diatoms. The process is most productive when the absorption of quanta are equal in both the PSII and PSI, assuring that input energy from the antenna complex is divided between the PSI and PSII system, which in turn powers the photochemistry.
Robert Hill thought that a complex of reactions consisting of an intermediate to cytochrome b6 (now a plastoquinone), another is from cytochrome f to a step in the carbohydrate-generating mechanisms. These are linked by plastoquinone, which does require energy to reduce cytochrome f for it is a sufficient reductant. Further experiments to prove that the oxygen developed during the photosynthesis of green plants came from water, were performed by Hill in 1937 and 1939. He showed that isolated chloroplasts give off oxygen in the presence of unnatural reducing agents like iron oxalate, ferricyanide or benzoquinone after exposure to light. The Hill reaction is as follows:
2 H2O + 2 A + (light, chloroplasts) → 2 AH2 + O2where A is the electron acceptor. Therefore, in light, the electron acceptor is reduced and oxygen is evolved.
Samuel Ruben and Martin Kamen used radioactive isotopes to determine that the oxygen liberated in photosynthesis came from the water.
Melvin Calvin and Andrew Benson, along with James Bassham, elucidated the path of carbon assimilation (the photosynthetic carbon reduction cycle) in plants. The carbon reduction cycle is known as the Calvin cycle, which ignores the contribution of Bassham and Benson. Many scientists refer to the cycle as the Calvin-Benson Cycle, Benson-Calvin, and some even call it the Calvin-Benson-Bassham (or CBB) Cycle.
Nobel Prize-winning scientist Rudolph A. Marcus was able to discover the function and significance of the electron transport chain.
Otto Heinrich Warburg and Dean Burk discovered the I-quantum photosynthesis reaction that splits the CO2, activated by the respiration.
Louis N.M. Duysens and Jan Amesz discovered that chlorophyll a will absorb one light, oxidize cytochrome f, chlorophyll a (and other pigments) will absorb another light, but will reduce this same oxidized cytochrome, stating the two light reactions are in series.
Development of the concept
In 1893, Charles Reid Barnes proposed two terms, photosyntax and photosynthesis, for the biological process of synthesis of complex carbon compounds out of carbonic acid, in the presence of chlorophyll, under the influence of light. Over time, the term photosynthesis came into common usage as the term of choice. Later discovery of anoxygenic photosynthetic bacteria and photophosphorylation necessitated redefinition of the term.
C3 : C4 photosynthesis research
After WWII at late 1940 at the University of California, Berkeley, the details of photosynthetic carbon metabolism were sorted out by the chemists Melvin Calvin, Andrew Benson, James Bassham and a score of students and researchers utilizing the carbon-14 isotope and paper chromatography techniques. The pathway of CO2 fixation by the algae Chlorella in a fraction of a second in light resulted in a 3 carbon molecule called phosphoglyceric acid (PGA). For that original and ground-breaking work, a Nobel Prize in Chemistry was awarded to Melvin Calvin 1961. In parallel, plant physiologists studied leaf gas exchanges using the new method of infrared gas analysis and a leaf chamber where the net photosynthetic rates ranged from 10 to 13 u mole CO2/square metere.sec., with the conclusion that all terrestrial plants having the same photosynthetic capacities that were light saturated at less than 50% of sunlight. These rates were determined in potted plants grown indoors under low light intensity.
Later in 1958-1963 at Cornell University, field grown maize was reported to have much greater leaf photosynthetic rates of 40 u mol CO2/square meter.sec and was not saturated at near full sunlight. This higher rate in maize was almost double those observed in other species such as wheat and soybean, indicating that large differences in photosynthesis exist among higher plants. At the University of Arizona, detailed gas exchange research on more than 15 species of monocot and dicot uncovered for the first time that differences in leaf anatomy are crucial factors in differentiating photosynthetic capacities among species. In tropical grasses, including maize, sorghum, sugarcane, Bermuda grass and in the dicot amaranthus, leaf photosynthetic rates were around 38−40 u mol CO2/square meter.sec., and the leaves have two types of green cells, i. e. outer layer of mesophyll cells surrounding a tightly packed cholorophyllous vascular bundle sheath cells. This type of anatomy was termed Kranz anatomy in the 19th century by the botanist Gottlieb Haberlandt while studying leaf anatomy of sugarcane. Plant species with the greatest photosynthetic rates and Kranz anatomy showed no apparent photorespiration, very low CO2 compensation point, high optimum temperature, high stomatal resistances and lower mesophyll resistances for gas diffusion and rates never saturated at full sun light. The research at Arizona was designated Citation Classic by the ISI 1986. These species was later termed C4 plants as the first stable compound of CO2 fixation in light has 4 carbon as malate and aspartate. Other species that lack Kranz anatomy were termed C3 type such as cotton and sunflower, as the first stable carbon compound is the 3-carbon PGA acid. At 1000 ppm CO2 in measuring air, both the C3 and C4 plants had similar leaf photosynthetic rates around 60 u mole CO2/square meter.sec. indicating the suppression of phototorespiration in C3 plants.
Factors
There are three main factors affecting photosynthesis and several corollary factors. The three main are:
Light intensity (irradiance), wavelength and temperature
The process of photosynthesis provides the main input of free energy into the biosphere, and is one of four main ways in which radiation is important for plant life.
The radiation climate within plant communities is extremely variable, with both time and space.
In the early 20th century, Frederick Blackman and Gabrielle Matthaei investigated the effects of light intensity (irradiance) and temperature on the rate of carbon assimilation.
These two experiments illustrate several important points: First, it is known that, in general, photochemical reactions are not affected by temperature. However, these experiments clearly show that temperature affects the rate of carbon assimilation, so there must be two sets of reactions in the full process of carbon assimilation. These are, of course, the light-dependent 'photochemical' temperature-independent stage, and the light-independent, temperature-dependent stage. Second, Blackman's experiments illustrate the concept of limiting factors. Another limiting factor is the wavelength of light. Cyanobacteria, which reside several meters underwater, cannot receive the correct wavelengths required to cause photoinduced charge separation in conventional photosynthetic pigments. To combat this problem, a series of proteins with different pigments surround the reaction center. This unit is called a phycobilisome.
Carbon dioxide levels and photorespiration
As carbon dioxide concentrations rise, the rate at which sugars are made by the light-independent reactions increases until limited by other factors. RuBisCO, the enzyme that captures carbon dioxide in the light-independent reactions, has a binding affinity for both carbon dioxide and oxygen. When the concentration of carbon dioxide is high, RuBisCO will fix carbon dioxide. However, if the carbon dioxide concentration is low, RuBisCO will bind oxygen instead of carbon dioxide. This process, called photorespiration, uses energy, but does not produce sugars.
RuBisCO oxygenase activity is disadvantageous to plants for several reasons:
- One product of oxygenase activity is phosphoglycolate (2 carbon) instead of 3-phosphoglycerate (3 carbon). Phosphoglycolate cannot be metabolized by the Calvin-Benson cycle and represents carbon lost from the cycle. A high oxygenase activity, therefore, drains the sugars that are required to recycle ribulose 5-bisphosphate and for the continuation of the Calvin-Benson cycle.
- Phosphoglycolate is quickly metabolized to glycolate that is toxic to a plant at a high concentration; it inhibits photosynthesis.
- Salvaging glycolate is an energetically expensive process that uses the glycolate pathway, and only 75% of the carbon is returned to the Calvin-Benson cycle as 3-phosphoglycerate. The reactions also produce ammonia (NH3), which is able to diffuse out of the plant, leading to a loss of nitrogen.
The salvaging pathway for the products of RuBisCO oxygenase activity is more commonly known as photorespiration, since it is characterized by light-dependent oxygen consumption and the release of carbon dioxide.