
 | ||
Homeostasis or homoeostasis is the property of a system within the body of a living organism in which a variable, such as the concentration of a substance in solution, is actively regulated to remain very nearly constant. Examples of homeostasis include the regulation of the body temperature of an animal, the pH of its extracellular fluids, or the concentrations of sodium (Na+) and calcium (Ca2+) ions or of glucose in the blood plasma, despite changes in the animal’s environment, or what it has eaten, or what it is doing (for example, resting or exercising). Each of these variables is controlled by a separate “homeostat” (or regulator), which, together, maintain life. Homeostats are energy-consuming physiological mechanisms.
Contents
- Etymology
- History
- Overview
- Core body temperature
- Blood glucose
- Plasma ionized calcium
- Blood partial pressure of oxygen and carbon dioxide
- Blood oxygen content
- Arterial blood pressure
- Extracellular sodium concentration
- Extracellular potassium concentration
- Volume of body water
- Extracellular fluid pH
- Homeostatic breakdown
- Chronic disease compensation and decompensation
- Examples from technology
- Biosphere
- Predictive
- Other fields
- Risk
- Stress
- References
The concept was described by French physiologist Claude Bernard in 1865 and the word was coined by Walter Bradford Cannon in 1926.
The concept of homeostasis is sometimes also applied to technological control systems such as thermostats, which function in an analogous way.
Etymology
The word homeostasis (/ˌhoʊmioʊˈsteɪsᵻs/) uses combining forms of homeo- and -stasis, New Latin from Greek: ὅμοιος homoios, "similar" and στάσις stasis, "standing still", yielding the idea of "staying the same".
History
The conceptual origins of homeostasis reach back to the ancient Greek concepts of balance, harmony, equilibrium, and steady-state, all of them fundamental attributes of life and health. Thus, the philosopher Empedocles (495-435 BC) postulated that all matter consisted of elements and qualities that were in dynamic opposition or alliance to one another, and that balance or harmony was a necessary condition for the survival of living organisms. Following these hypotheses, Hippocrates (460-375 BC) compared health to the harmonious balance of the elements, and illness and disease to the systematic disharmony of these elements.
Nearly 150 years ago, the French physiologist Claude Bernard published his seminal work, stating that the maintenance of the internal environment, or milieu intérieur, surrounding the body's cells, was essential for the life of the organism. In 1929, Walter B. Cannon published an extrapolation from Bernard's 1865 work naming his theory "homeostasis". Cannon postulated that homeostasis was a process of synchronized adjustments in the internal environment resulting in the maintenance of specific physiological variables within defined parameters; and that these precise parameters included blood pressure, temperature, pH, and others; all with clearly defined "normal" ranges. Cannon further posited that threats to homeostasis might originate from the external environment (e.g., temperature extremes, traumatic injury) or the internal environment (e.g., pain, infection), and could be physical or psychological, as in emotional distress. Cannon's work outlined that maintenance of this internal physical and psychological balance, homeostasis, demands an internal network of communication, with sensors capable of identifying deviations from the acceptable ranges and effectors to return those deviations back within acceptable limits. Cannon identified these negative feedback systems and emphasized that, regardless of the nature of the threat to homeostasis, the response he mapped within the body would be the same.
Overview
The metabolic processes of all living organisms can only take place in very specific physical and chemical environments. The conditions vary with each organism, and with whether the chemical processes take place inside the cell or in the fluids bathing the cells in multicellular creatures. The best known homeostats in human and other mammalian bodies are regulators that keep the composition of the extracellular fluids (or the ”internal environment”) constant, especially with regard to the temperature, pH, osmolality, and the concentrations of Na+, K+, Ca2+, glucose and CO2 and O2. However, a great many other homeostats, encompassing many aspects of human physiology, control other entities in the body.
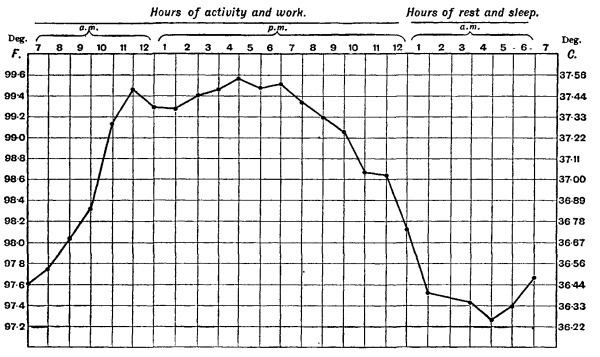
If an entity is homeostatically controlled it does not imply that its value is necessarily absolutely steady in health. Core body temperature is, for instance, regulated by a homeostat with temperature sensors in, amongst others, the hypothalamus of the brain. However the set point of the regulator is regularly reset. For instance, core body temperature in humans varies during the course of the day (i.e. has a circadian rhythm), with the lowest temperatures occurring at night, and the highest in the afternoons (see diagram on the right). The temperature regulator's set point is readjusted in adult women at the start of the luteal phase of the menstrual cycle (see the diagram on the right, below). The temperature regulator's set point is reset during infections to produce a fever.
Homeostasis does not govern every activity in the body. For instance the signal (be it via neurons or hormones) from the sensor to the effector is, of necessity, highly variable in order to convey information about the direction and magnitude of the error detected by the sensor. Similarly the effector’s response needs to be highly adjustable to reverse the error – in fact it should be very nearly in proportion (but in the opposite direction) to the error that is threatening the internal environment. For instance, the arterial blood pressure in mammals is homeostatically controlled, and measured by sensors in the aorta and carotid arteries. The sensors send messages via sensory nerves to the medulla oblongata of the brain indicating whether the blood pressure has fallen or risen, and by how much. The medulla oblongata then distributes messages along motor or efferent nerves belonging to the autonomic nervous system to a wide variety of effector organs, whose activity is consequently changed to reverse the error in the blood pressure. One of the effector organs is the heart whose rate is stimulated to rise (tachycardia) when the arterial blood pressure falls, or to slow down (bradycardia) when the pressure rises above set point. Thus the heart rate (for which there is no sensor in the body) is not homeostatically controlled, but is one of effector responses to errors in the arterial blood pressure. Another example is the rate of sweating. This is one of the effectors in the homeostatic control of body temperature, and therefore highly variable in rough proportion to the heat load that threatens to destabilize the body’s core temperature, for which there is a sensor in the hypothalamus of the brain.
Apart from the entities that are homeostatically controlled in the internal environment of the body, and the mechanisms that are responsible for this regulation, there are variables that are neither homeostatically controlled nor involved in the operation of homeostats. The blood urea concentration is an example. Mammals do not have "urea sensors". Instead the concentration of urea is determined by a dynamic equilibrium, in much the same way that the water level in a river at any particular point along its course is determined. The level of a river is simply dependent on the rate at which water flows into a particular section and how fast it flows away from there. It therefore varies with the rainfall in the catchment area and obstructions or otherwise to the flow down stream – there is no energy consuming “regulation”. The blood urea concentration is comparable to the water level in a natural river. It is manufactured by the liver from the amino groups of the amino acids of proteins that are being degraded in this organ. It is then excreted by the kidneys which simply pass most of the urea in the glomerular filtrate on into the urine without active resorption or excretion by the renal tubules (a relatively small proportion of the urea in the tubules diffuses passively back into the blood as its concentration in the tubules rises when water, without urea, is removed from the tubular fluid). A high protein diet therefore produces high blood urea concentrations, and a protein-poor diet produced low blood plasma urea concentrations, without any physiological attempt to correct or mitigate these fluctuations in the level of urea in the extracellular fluids.
Core body temperature
Mammals regulate their core temperatures, using hypothalamic temperature sensors in their brains, but also elsewhere in their bodies. When core body temperature falls behavioral changes are set in motion, which, in humans, include the donning of warmer clothes, the seeking out of wind-free, warmer environments, and, eventually, the curling up in the "fetal position" to reduce the surface area (skin) exposed to the cold.
The metabolic rate is increased, initially by non-shivering thermogenesis, followed by shivering thermogenesis if the earlier reactions are insufficient to correct the hypothermia.
When body temperature rises, or skin heat sensors detect a threatening rise in body temperature, behavioral changes cause the animal to seek shade, and, in humans, the sweat glands in the skin are stimulated via cholinergic sympathetic nerves to secrete a dilute watery fluid called sweat onto the skin, which, when it evaporates, cools the skin and the blood flowing through it. Panting is an alternative effector in many vertebrates, which cools the body also by the evaporation of water, but this time from the mucous membranes of the throat and mouth.
Blood glucose
All animals regulate the glucose concentration in their extracellular fluids. In mammals the primary sensor is situated in the beta cells of the pancreatic islets. The beta cells respond to a rise in the blood sugar level by secreting insulin into the blood, and simultaneously inhibiting their neighboring alpha cells from secreting glucagon into the blood. This combination (high blood insulin levels and low glucagon levels) act on effector tissues, chief of which are the liver, fat cells and muscle cells. The liver is inhibited from producing glucose, taking it up instead, and converting it to glycogen and triglycerides. The glycogen is stored in the liver, but the triglycerides are secreted into the blood as very low-density lipoprotein (VLDL) particles which are taken up by adipose tissue, there to be stored as fats. The fat cells take up glucose through special glucose transporters (GLUT4), whose numbers in the cell wall are increased as a direct effect of insulin acting on these cells. The glucose that enters the fat cells in this manner is converted into triglycerides (via the same metabolic pathways as are used by the liver) and then stored in those fat cells together with the VLDL-derived triglycerides that were made in the liver. Muscle cells also take glucose up through insulin-sensitive GLUT4 glucose channels, and convert it into muscle glycogen.
When the beta cells in the pancreatic islets detect lower than normal blood glucose levels, insulin secretion into the blood ceases and the alpha cells are stimulated to secrete glucagon into the blood. This inhibits the uptake of glucose from the blood by the liver, fats cells and muscle. Instead the liver is strongly stimulated to manufacture glucose from glycogen (through glycogenolysis) and from non-carbohydrate sources (such as lactate and de-aminated amino acids) using a process known as gluconeogenesis. The glucose thus produced is discharged into the blood correcting the detected error (hypoglycemia). The glycogen stored in muscles remains in the muscles, and is only broken down, during exercise, to glucose-6-phosphate and thence to pyruvate to be fed into the citric acid cycle or turned into lactate. It is only the lactate and the waste products of the citric acid cycle that are returned to the blood. The liver can take up only the lactate, and by the process of energy consuming gluconeogenesis convert it back to glucose.
Plasma ionized calcium
The plasma ionized calcium (Ca2+) concentration is very tightly controlled by a pair of homeostats. The sensor for the one is situated in the parathyroid glands, where the chief cells sense the Ca2+ level by means of specialized calcium receptors in their membranes. The sensors for the second homeostat are the parafollicular cells in the thyroid gland. The parathyroid chief cells secrete parathyroid hormone (PTH) in response to a fall in the plasma ionized calcium level; the parafollicular cells of the thyroid gland secrete calcitonin in response to a rise in the plasma ionized calcium level.
The effector organs of the first homeostat are the skeleton, the kidney, and, via a hormone released into the blood by the kidney in response to high PTH levels in the blood, the duodenum and jejunum. Parathyroid hormone (in high concentrations in the blood) causes bone resorption, releasing calcium into the plasma. This is a very rapid action which can correct a threatening hypocalcemia within minutes. High PTH concentrations cause the excretion of phosphate ions via the urine. Since phosphates combine with calcium ions to form insoluble salts, a decrease in the level of phosphates in the blood, releases free calcium ions into the plasma ionized calcium pool. PTH has a second action on the kidneys. It stimulates the manufacture and release, by the kidneys, of calcitriol (or 1,25 dihydroxycholecalciferol, or 1,25 dihydroxyvitamin D3) into the blood. This steroid hormone acts on the epithelial cells of the upper small intestine, increasing their capacity to absorb calcium from the gut contents into the blood.
The second homeostat, with its sensors in the thyroid gland, releases calcitonin into the blood when the blood ionized calcium rises. This hormone acts primarily on bone, causing the rapid removal of calcium from the blood and depositing it, in insoluble form, in the skeleton.
The two homeostats working through PTH on the one hand, and calcitonin on the other, can very rapidly correct any impending error in the plasma ionized calcium level by either removing calcium from the blood and depositing it in the skeleton, or by removing calcium from it. The skeleton acts as an extremely large calcium store (about 1 kg) compared with the plasma calcium store (about 180 mg). Longer term regulation occurs through calcium absorption or loss from the gut (see Regulation of calcium metabolism in the Calcium metabolism article).
Blood partial pressure of oxygen and carbon dioxide
The partial pressure of oxygen (
Too little CO2, and, to a lesser extent, too much O2, in the blood can temporarily halt breathing, which breath-holding divers use to prolong the time they can stay underwater.
The
3) into the urine. This is an important contribution to the acclimatization to high altitude.
Blood oxygen content
The kidneys measure the oxygen content (rather than the
The distinction between the
Arterial blood pressure
Stretch receptors in the walls of the aortic arch and carotid sinus (at the beginning of the internal carotid artery) act as arterial blood pressure sensors. As the pressure rises the arteries balloon out, stretching their walls. This information is then conveyed, via sensory nerves, to the medulla oblongata of the brain stem. From here motor nerves belonging to the autonomic nervous system are stimulated to influence the activity of chiefly the heart and the smallest diameter arteries, called arterioles. The arterioles are the main resistance vessels in the arterial tree, and small changes in diameter cause large changes in the resistance to flow through them. When the arterial blood pressure rises the arterioles are stimulated to dilate making it easier for blood to leave the arteries, thus deflating them, and bringing the blood pressure down, back to normal. At the same time the heart is stimulated via cholinergic parasympathetic nerves to beat more slowly (called bradycardia), ensuring that the inflow of blood into the arteries is reduced, thus adding to the reduction in pressure, and correction of the original error.
If the pressure in the arteries falls, the opposite reflex is elicited: constriction of the arterioles, and a speeding up of the heart rate (called tachycardia). If the drop in blood pressure is very rapid or excessive, the medulla oblongata stimulates the adrenal medulla, via "preganglionic" sympathetic nerves, to secrete epinephrine (adrenaline) into the blood. This hormone enhances the tachycardia and causes severe vasoconstriction of the arterioles to all but the essential organ in the body (especially the heart, lungs and brain). These reactions usually correct the low arterial blood pressure (hypotension) very effectively.
Extracellular sodium concentration
The sodium concentration homeostat is rather more complex than most of the other homeostats described on this page.
The sensor is situated in the juxtaglomerular apparatus of kidneys, which senses the plasma sodium concentration in a surprisingly indirect manner. Instead of measuring it directly in the blood flowing past the juxtaglomerular cells, these cells respond to the sodium concentration in the renal tubular fluid after it has already undergone a certain amount of modification in the proximal convoluted tubule and loop of Henle. These cells also respond to rate of blood flow through the juxtaglomerular apparatus, which, under normal circumstances, is directly proportional to the arterial blood pressure, making this tissue an ancillary arterial blood pressure sensor.
In response to a lowering of the plasma sodium concentration, or to a fall in the arterial blood pressure, the juxtaglomerular cells release renin into the blood. Renin is an enzyme which cleaves a decapeptide (a short protein chain, 10 amino acids long) from a plasma α-2-globulin called angiotensinogen. This decapeptide is known as angiotensin I. It has no known biological activity. However, when the blood circulates through the lungs a pulmonary capillary endothelial enzyme called angiotensin-converting enzyme (ACE) cleaves a further two amino acids from angiotensin I to form an octapeptide known as angiotensin II. Angiotensin II is a hormone which acts on the adrenal cortex, causing the release into the blood of the steroid hormone, aldosterone. Angiotensin II also acts on the smooth muscle in the walls of the arterioles causing these small diameter vessels to constrict, thereby restricting the outflow of blood from the arterial tree, causing the arterial blood pressure to rise. This therefore reinforces the measures described above (under the heading of The arterial blood pressure homeostat), which defend the arterial blood pressure against changes, especially hypotension.
The angiotensin II-stimulated aldosterone released from the zona glomerulosa of the adrenal glands has an effect on particularly the epithelial cells of the distal convoluted tubules and collecting ducts of the kidneys. Here it causes the reabsorption of sodium ions from the renal tubular fluid, in exchange for potassium ions which are secreted from the blood plasma into the tubular fluid to exit the body via the urine. The reabsorption of sodium ions from the renal tubular fluid halts further sodium ion losses from the body, and therefore preventing the worsening of hyponatremia. The hyponatremia can only be corrected by the consumption of salt in the diet. However, it is not certain whether a “salt hunger” can be initiated by hyponatremia, or by what mechanism this might come about.
When the plasma sodium ion concentration is higher than normal (hypernatremia), the release of renin from the juxtaglomerular apparatus is halted, ceasing the production of angiotensin II, and its consequent aldosterone-release into the blood. The kidneys respond by excreting sodium ions into the urine, thereby normalizing the plasma sodium ion concentration. The low angiotensin II levels in the blood lower the arterial blood pressure as an inevitable concomitant response.
The reabsorption of sodium ions from the tubular fluid as a result of high aldosterone levels in the blood does not, of itself, cause renal tubular water to be returned to the blood from the distal convoluted tubules or collecting ducts. This is because sodium is reabsorbed in exchange for potassium and therefore causes only a modest change in the osmotic gradient between the blood and the tubular fluid. Furthermore, the epithelium of the distal convoluted tubules and collecting ducts is impermeable to water in the absence of antidiuretic hormone (ADH) in the blood. ADH is part of the body water homeostat. Its levels in the blood vary with the osmolality of the plasma, which is measured in the hypothalamus of the brain. Aldosterone's action on the kidney tubules does not add sodium to the extracellular fluids (ECF) - it simply prevents further loss. So there is no change in the osmolality of the ECF, and therefore no change in the ADH concentration of the plasma. However, low aldosterone levels cause a loss of sodium ions from the ECF, which could potentially cause a change in extracellular osmolality and therefore of ADH levels in the blood.
Extracellular potassium concentration
The extracellular potassium ion (K+) concentration is sensed by the zona glomerulosa cells of the outer layer of the adrenal cortex, as well as, probably, by sensors in the carotid arteries. High potassium concentrations in the plasma cause depolarization of the zona glomerulosa cells’ membranes. This causes the release of aldosterone into the blood.
Aldosterone acts primarily on the distal convoluted tubules and collecting ducts of the kidneys, stimulating them to excrete potassium ions into the tubular fluid, and thus into the urine. It does so, however, by activating the basolateral Na+/K+ pumps of the tubular epithelial cells. These sodium/potassium exchangers pump three sodium ions out of the cell, into the interstitial fluid and two potassium ions into the cell from the interstitial fluid. This creates concentration gradients which result in the reabsorption of sodium (Na+) ions from the tubular fluid into the blood, and secreting potassium (K+) ions from the blood into the urine (lumen of collecting duct).
This obviously implies that excess potassium in the plasma can only be excreted at the expense of sodium retention by the body. The fact that the sodium and potassium homeostats seem to rely entirely on the same effector, but in opposite directions, implies that the body can only excrete potassium while retaining sodium, or vice versa. It cannot simultaneously excrete sodium and potassium ions in higher than modest quantities (when aldosterone is at an intermediate concentration in the plasma), and has no way of retaining both of them if there is a shortage in the body of the two cations. How these two conflicting homeostats (using the same effector) are disentangled to allow the plasma sodium and potassium ion levels to be regulated independently is currently not clear.
Volume of body water
The volume of water in the body is measured by stretch receptors in the heart atria, and, somewhat indirectly, by the measurement of the osmolality of the plasma by the hypothalamus. Measurement of the plasma osmolality to give an indication of the water content of the body, relies on the fact that water losses from the body, through sweat, gut fluids (normal fecal water losses, and through vomiting and diarrhea), and the exhaled air, are all hypotonic, meaning that they are less salty than the body fluids (compare, for instance, the taste of saliva with that of tears. The latter have almost the same salt content as the extracellular fluids, whereas the former is hypotonic with respect to plasma. Saliva does not taste salty, whereas tears are decidedly salty). Nearly all normal and abnormal losses of body water therefore cause the extracellular fluids to become hyperosmolar. Conversely excessive water intake (in the form of most regular beverages) dilutes the extracellular fluids causing the hypothalamus to register hypo-osmolar conditions.
When the hypothalamus detects a hyperosmolar extracellular environment, it causes the secretion from the posterior pituitary gland of a peptide hormone called antidiuretic hormone (ADH), which acts on the effector organ, which in this case is the kidney. The effect of ADH on the kidney tubules is to reabsorb water from the distal convoluted tubules and collecting ducts, thus preventing aggravation of the water loss via the urine. The hypothalamus simultaneously stimulates the nearby thirst center causing an almost irresistible (if the hyperosmolarity is severe enough) urge to drink water. The cessation of urine flow prevents the hypovolemia and hypertonicity from getting worse; the drinking of water corrects the defect.
Hypo-osmolality results in very low plasma ADH levels. This results in the inhibition of water reabsorption from the kidney tubules, causing high volumes of very dilute urine to be excreted, thus getting rid of the excess water in the body.
Note that urinary water loss, when the body water homeostat is intact, is a compensatory water loss, correcting any water excess in the body. However, since the kidneys cannot generate water, the thirst reflex is the all important second effector mechanism of the body water homeostat, correcting any water deficit in the body.
Stretching of the right atrium of the heart, usually a sign of an excessive blood volume, causes stretch receptors to secrete a hormone known as atrial natriuretic peptide (ANP) into the blood. This also acts on the kidneys causing sodium, and accompanying water loss into the urine, thereby reducing the volume of circulating blood.
Extracellular fluid pH
The pH of the extracellular fluids (which includes the blood plasma) is regulated by adjusting the ratio of the concentration of carbonic acid (H2CO3) to that of the bicarbonate ions (HCO−
3) to equal 1:20. This ratio and its relationship to the pH is described by the Henderson–Hasselbalch equation, which, when applied to the bicarbonate buffering system in the extracellular fluids, states that:
where:
3] is the concentration of bicarbonate in the blood plasma
However, since the carbonic acid concentration is directly proportional to the
where:
3] is the concentration of bicarbonate in the plasma
There are therefore at least two homeostats responsible for the regulation of the plasma pH. The first is the
The sensor for the plasma HCO−
3 concentration is not known for certain. It is very probable that the renal tubular cells of the distal convoluted tubules are themselves sensitive to the pH of the plasma. The metabolism of these cells produces CO2, which is rapidly converted to H+ and HCO−
3 through the action of carbonic anhydrase. When the extracellular fluids tend towards acidity, the renal tubular cells secrete the H+ ions into the tubular fluid from where they exit the body via the urine. The HCO−
3 ions are simultaneously secreted into the blood plasma, thus raising the bicarbonate ion concentration in the plasma, increasing the [
Homeostatic breakdown
Many diseases are the result of the failure of one or more homeostat(s). Almost any functional component of any homeostat can malfunction, either as a result of an inherited defect, or an acquired disease. Some of the homeostats have inbuilt redundancies, which insures that life is not immediately threatened if a component malfunctions; but in other cases malfunction of a homeostat causes severe disease, which can be fatal if not treated. Here only a few well known examples of homeostat dysfunction are described.
Type 1 diabetes mellitus is probably the best known example. Here the blood glucose homeostat ceases to function because the beta cells of the pancreatic islets are destroyed. This means that the glucose sensor is absent, and its effector pathway (the insulin level in the blood) remains unchanged at zero. The blood glucose concentration therefore rises to very high levels, while the body’s proteins are degraded into amino acids which are turned at a very high rate into glucose, via gluconeogenesis, by the liver. The condition is fatal if not treated.
The plasma ionized calcium homeostat can be disrupted by the constant, unchanging, over-production of parathyroid hormone by a parathyroid adenoma resulting in the typically features of hyperparathyroidism, namely high plasma ionized Ca2+ levels and the resorption of bone, which can lead to spontaneous fractures. The abnormally high plasma ionized calcium concentrations cause conformational changes in many cell-surface proteins (especially ion channels and hormone or neurotransmitter receptors) giving rise to lethargy, muscle weakness, anorexia, constipation and labile emotions.
The body water homeostat can be compromised by the inability to secrete ADH in response to even the normal daily water losses via the exhaled air, the feces, and insensible sweating. On receiving a zero blood ADH signal, the kidneys produce huge unchanging volumes of very dilute urine, causing dehydration and death if not treated.
As organisms age, the efficiency of their control systems becomes reduced. The inefficiencies gradually result in an unstable internal environment that increases the risk of illness, and leads to the physical changes associated with aging.
Chronic disease compensation and decompensation
Various chronic diseases are kept under control by homeostatic compensation, which masks a problem by compensating for it (making up for it) in another way. However, the compensating mechanisms eventually wear out or are disrupted by a new complicating factor (such as the advent of a concurrent acute viral infection), which sends the body reeling through a new cascade of events. Such decompensation unmasks the underlying disease, worsening its symptoms. Common examples include decompensated heart failure, kidney failure, and liver failure.
Examples from technology
Familiar technological homeostatic mechanisms include:
Biosphere
In the Gaia hypothesis, James Lovelock stated that the entire mass of living matter on Earth (or any planet with life) functions as a vast homeostatic superorganism that actively modifies its planetary environment to produce the environmental conditions necessary for its own survival. In this view, the entire planet maintains several homeostats (the primary one being temperature homeostasis). Whether this sort of system is present on Earth is open to debate. However, some relatively simple homeostatic mechanisms are generally accepted. For example, it is sometimes claimed that when atmospheric carbon dioxide levels rise, certain plants may be able to grow better and thus act to remove more carbon dioxide from the atmosphere. However, warming has exacerbated droughts, making water the actual limiting factor on land. When sunlight is plentiful and atmospheric temperature climbs, it has been claimed that the phytoplankton of the ocean surface waters, acting as global sunshine, and therefore heat sensors, may thrive and produce more dimethyl sulfide (DMS). The DMS molecules act as cloud condensation nuclei, which produce more clouds, and thus increase the atmospheric albedo, and this feeds back to lower the temperature of the atmosphere. However, rising sea temperature has stratified the oceans, separating warm, sunlit waters from cool, nutrient-rich waters. Thus, nutrients have become the limiting factor, and plankton levels have actually fallen over the past 50 years, not risen. As scientists discover more about Earth, vast numbers of positive and negative feedback loops are being discovered, that, together, maintain a metastable condition, sometimes within very broad range of environmental conditions.
Predictive
Predictive homeostasis is an anticipatory response to an expected challenge in the future, such as the stimulation of insulin secretion by gut hormones which enter the blood in response to a meal. This insulin secretion occurs before the blood sugar level rises, lowering the blood sugar level in anticipation of a large influx into the blood of glucose resulting from the digestion of carbohydrates in the gut. Such anticipatory reactions are open loop systems which are based, essentially, on “guess work”, and are not self-correcting. Anticipatory responses always require a closed loop negative feedback system to correct the over- and undershoots to which the anticipatory systems are prone.
Other fields
The term has come to be used in other fields, for example:
Risk
An actuary may refer to risk homeostasis, where (for example) people who have anti-lock brakes have no better safety record than those without anti-lock brakes, because the former unconsciously compensate for the safer vehicle via less-safe driving habits. Previous to the innovation of anti-lock brakes, certain maneuvers involved minor skids, evoking fear and avoidance: Now the anti-lock system moves the boundary for such feedback, and behavior patterns expand into the no-longer punitive area. It has also been suggested that ecological crises are an instance of risk homeostasis in which a particular behavior continues until proven dangerous or dramatic consequences actually occur.
Stress
Sociologists and psychologists may refer to stress homeostasis, the tendency of a population or an individual to stay at a certain level of stress, often generating artificial stresses if the "natural" level of stress is not enough.
Jean-François Lyotard, a postmodern theorist, has applied this term to societal 'power centers' that he describes as being 'governed by a principle of homeostasis,' for example, the scientific hierarchy, which will sometimes ignore a radical new discovery for years because it destabilises previously accepted norms. (See The Postmodern Condition: A Report on Knowledge by Jean-François Lyotard)