
 | ||
Insect morphology is the study and description of the physical form of insects. The terminology used to describe insects is similar to that used for other arthropods due to their shared evolutionary history. Three physical features separate insects from other arthropods: they have a body divided into three regions (head, thorax, and abdomen), have three pairs of legs, and mouthparts located outside of the head capsule. It is this position of the mouthparts which divides them from their closest relatives, the non-insect hexapods, which includes Protura, Diplura, and Collembola.
Contents
- Anatomy summary
- Nervous system
- Digestive system
- Foregut
- Midgut
- Hindgut
- Respiratory systems
- Circulatory system
- Endocrine system
- Female
- Male
- Blattodea
- Coleoptera
- Dermaptera
- Diptera
- Lepidoptera
- References
There is enormous variation in body structure amongst insect species. Individuals can range from 0.3 mm (fairyflies) to 30 cm across (great owlet moth); have no eyes or many; well-developed wings or none; and legs modified for running, jumping, swimming, or even digging. These modifications allow insects to occupy almost every ecological niche on the planet, except the deep ocean and the Antarctic. This article describes the basic insect body and some of the major variations of the different body parts; in the process it defines many of the technical terms used to describe insect bodies.
Anatomy summary
Insects, like all arthropods, have no interior skeleton; instead, they have an exoskeleton, a hard outer layer made mostly of chitin which protects and supports the body. The insect body is divided into three parts: the head, thorax, and abdomen. The head is specialized for sensory input and food intake; the thorax, which is the anchor point for the legs and wings (if present), is specialized for locomotion; and the abdomen for digestion, respiration, excretion, and reproduction. Although the general function of the three body regions is the same across all insect species, there are major differences in basic structure, with wings, legs, antennae, and mouthparts being highly variable from group to group.
Nervous system
The nervous system of an insect can be divided into a brain and a ventral nerve cord. The head capsule is made up of six fused segments, each with a pair of ganglia, or a cluster of nerve cells outside of the brain. The first three pairs of ganglia are fused into the brain, while the three following pairs are fused into a structure of three pairs of ganglia under the insect's esophagus, called the subesophageal ganglion.
The thoracic segments have one ganglion on each side, which are connected into a pair, one pair per segment. This arrangement is also seen in the abdomen but only in the first eight segments. Many species of insects have reduced numbers of ganglia due to fusion or reduction. Some cockroaches have just six ganglia in the abdomen, whereas the wasp Vespa crabro has only two in the thorax and three in the abdomen. Some insects, like the house fly Musca domestica, have all the body ganglia fused into a single large thoracic ganglion.
At least a few insects have nociceptors, cells that detect and transmit sensations of pain. This was discovered in 2003 by studying the variation in reactions of larvae of the common fruitfly Drosophila to the touch of a heated probe and an unheated one. The larvae reacted to the touch of the heated probe with a stereotypical rolling behavior that was not exhibited when the larvae were touched by the unheated probe. Although nociception has been demonstrated in insects, there is not a consensus that insects feel pain consciously.
Digestive system
An insect uses its digestive system to extract nutrients and other substances from the food it consumes. Most of this food is ingested in the form of macromolecules and other complex substances like proteins, polysaccharides, fats, and nucleic acids. These macromolecules must be broken down by catabolic reactions into smaller molecules like amino acids and simple sugars before being used by cells of the body for energy, growth, or reproduction. This break-down process is known as digestion. The main structure of an insect's digestive system is a long enclosed tube called the alimentary canal, which runs lengthwise through the body. The alimentary canal directs food in one direction: from the mouth to the anus. It has three sections, each of which performs a different process of digestion. In addition to the alimentary canal, insects also have paired salivary glands and salivary reservoirs. These structures usually reside in the thorax, adjacent to the foregut. The gut is where almost all of insects' digestion takes place. It can be divided into the foregut, midgut and hindgut.
Foregut
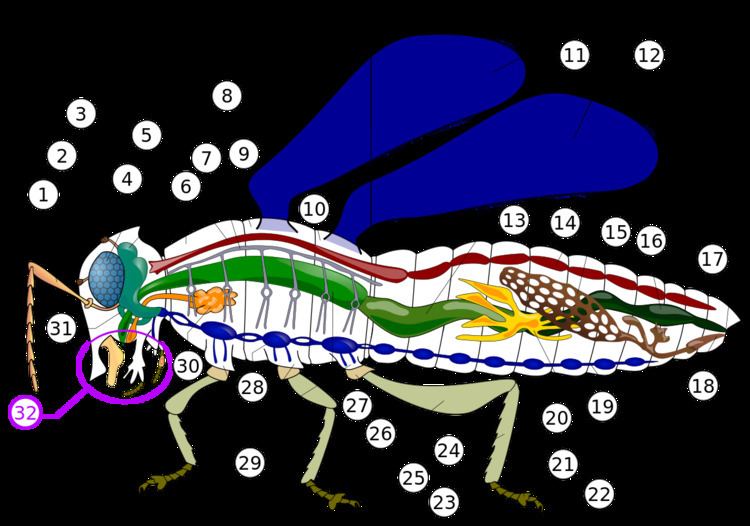
The first section of the alimentary canal is the foregut (element 27 in numbered diagram), or stomodaeum. The foregut is lined with a cuticular lining made of chitin and proteins as protection from tough food. The foregut includes the buccal cavity (mouth), pharynx, esophagus, and Crop and proventriculus (any part may be highly modified), which both store food and signify when to continue passing onward to the midgut. Here, digestion starts as partially chewed food is broken down by saliva from the salivary glands. As the salivary glands produce fluid and carbohydrate-digesting enzymes (mostly amylases), strong muscles in the pharynx pump fluid into the buccal cavity, lubricating the food like the salivarium does, and helping blood feeders, and xylem and phloem feeders.
From there, the pharynx passes food to the esophagus, which could be just a simple tube passing it on to the crop and proventriculus, and then on ward to the midgut, as in most insects. Alternately, the foregut may expand into a very enlarged crop and proventriculus, or the crop could just be a diverticulum, or fluid filled structure, as in some Diptera species.
The salivary glands (element 30 in numbered diagram) in an insect's mouth produce saliva. The salivary ducts lead from the glands to the reservoirs and then forward through the head to an opening called the salivarium, located behind the hypopharynx. By moving its mouthparts (element 32 in numbered diagram) the insect can mix its food with saliva. The mixture of saliva and food then travels through the salivary tubes into the mouth, where it begins to break down. Some insects, like flies, have extra-oral digestion. Insects using extra-oral digestion expel digestive enzymes onto their food to break it down. This strategy allows insects to extract a significant proportion of the available nutrients from the food source.
Midgut
Once food leaves the crop, it passes to the midgut (element 13 in numbered diagram), also known as the mesenteron, where the majority of digestion takes place. Microscopic projections from the midgut wall, called microvilli, increase the surface area of the wall and allow more nutrients to be absorbed; they tend to be close to the origin of the midgut. In some insects, the role of the microvilli and where they are located may vary. For example, specialized microvilli producing digestive enzymes may more likely be near the end of the midgut, and absorption near the origin or beginning of the midgut.
Hindgut
In the hindgut (element 16 in numbered diagram), or proctodaeum, undigested food particles are joined by uric acid to form fecal pellets. The rectum absorbs 90% of the water in these fecal pellets, and the dry pellet is then eliminated through the anus (element 17), completing the process of digestion. The uric acid is formed using hemolymph waste products diffused from the Malpighian tubules (element 20). It is then emptied directly into the alimentary canal, at the junction between the midgut and hindgut. The number of Malpighian tubules possessed by a given insect varies between species, ranging from only two tubules in some insects to over 100 tubules in others.
Respiratory systems
Insect respiration is accomplished without lungs. Instead, the insect respiratory system uses a system of internal tubes and sacs through which gases either diffuse or are actively pumped, delivering oxygen directly to tissues that need it via their trachea (element 8 in numbered diagram). Since oxygen is delivered directly, the circulatory system is not used to carry oxygen, and is therefore greatly reduced. The insect circulatory system has no veins or arteries, and instead consists of little more than a single, perforated dorsal tube that pulses peristaltically. Toward the thorax, the dorsal tube (element 14) divides into chambers and acts like the insect's heart. The opposite end of the dorsal tube is like the aorta of the insect circulating the hemolymph, arthropods' fluid analog of blood, inside the body cavity. Air is taken in through openings on the sides of the abdomen called spiracles.
There are many different patterns of gas exchange demonstrated by different groups of insects. Gas exchange patterns in insects can range from continuous and diffusive ventilation, to discontinuous gas exchange. During continuous gas exchange, oxygen is taken in and carbon dioxide is released in a continuous cycle. In discontinuous gas exchange, however, the insect takes in oxygen while it is active and small amounts of carbon dioxide are released when the insect is at rest. Diffusive ventilation is simply a form of continuous gas exchange that occurs by diffusion rather than physically taking in the oxygen. Some species of insect that are submerged also have adaptations to aid in respiration. As larvae, many insects have gills that can extract oxygen dissolved in water, while others need to rise to the water surface to replenish air supplies, which may be held or trapped in special structures.
Circulatory system
Insect blood or haemolymph's main function is that of transport and it bathes the insect's body organs. Making up usually less than 25% of an insect's body weight, it transports hormones, nutrients and wastes and has a role in, osmoregulation, temperature control, immunity, storage (water, carbohydrates and fats) and skeletal function. It also plays an essential part in the moulting process. An additional role of the haemolymph in some orders, can be that of predatory defence. It can contain unpalatable and malodourous chemicals that will act as a deterrent to predators. Haemolymph contains molecules, ions and cells; regulating chemical exchanges between tissues, haemolymph is encased in the insect body cavity or haemocoel. It is transported around the body by combined heart (posterior) and aorta (anterior) pulsations, which are located dorsally just under the surface of the body. It differs from vertebrate blood in that it doesn't contain any red blood cells and therefore is without high oxygen carrying capacity, and is more similar to lymph found in vertebrates.
Body fluids enter through one-way valved ostia, which are openings situated along the length of the combined aorta and heart organ. Pumping of the haemolymph occurs by waves of peristaltic contraction, originating at the body's posterior end, pumping forwards into the dorsal vessel, out via the aorta and then into the head where it flows out into the haemocoel. The haemolymph is circulated to the appendages unidirectionally with the aid of muscular pumps or accessory pulsatile organs usually found at the base of the antennae or wings and sometimes in the legs, with pumping rates accelerating with periods of increased activity. Movement of haemolymph is particularly important for thermoregulation in orders such as Odonata, Lepidoptera, Hymenoptera and Diptera.
Endocrine system
These glands are part of the endocrine system:
1. Neurosecretory cells
2. Corpora cardiaca
4. Corpora allata
Female
Female insects are able make eggs, receive and store sperm, manipulate sperm from different males, and lay eggs. Their reproductive systems are made up of a pair of ovaries, accessory glands, one or more spermathecae, and ducts connecting these parts. The ovaries make eggs and accessory glands produce the substances to help package and lay the eggs. Spermathecae store sperm for varying periods of time and, along with portions of the oviducts, can control sperm use. The ducts and spermathecae are lined with a cuticle.
The ovaries are made up of a number of egg tubes, called ovarioles, which vary in size and number by species. The number of eggs that the insect is able to make vary by the number of ovarioles with the rate that eggs can be develop being also influenced by ovariole design. In meroistic ovaries, the eggs-to-be divide repeatedly and most of the daughter cells become helper cells for a single oocyte in the cluster. In panoistic ovaries, each egg-to-be produced by stem germ cells develops into an oocyte; there are no helper cells from the germ line. Production of eggs by panoistic ovaries tends to be slower than that by meroistic ovaries.
Accessory glands or glandular parts of the oviducts produce a variety of substances for sperm maintenance, transport, and fertilization, as well as for protection of eggs. They can produce glue and protective substances for coating eggs or tough coverings for a batch of eggs called oothecae. Spermathecae are tubes or sacs in which sperm can be stored between the time of mating and the time an egg is fertilized. Paternity testing of insects has revealed that some, and probably many, female insects use the spermatheca and various ducts to control or bias sperm used in favor of some males over others.
Male
The main component of the male reproductive system is the testis, suspended in the body cavity by tracheae and the fat body. The more primitive apterygote insects have a single testis, and in some lepidopterans the two maturing testes are secondarily fused into one structure during the later stages of larval development, although the ducts leading from them remain separate. However, most male insects have a pair of testes, inside of which are sperm tubes or follicles that are enclosed within a membranous sac. The follicles connect to the vas deferens by the vas efferens, and the two tubular vasa deferentia connect to a median ejaculatory duct that leads to the outside. A portion of the vas deferens is often enlarged to form the seminal vesicle, which stores the sperm before they are discharged into the female. The seminal vesicles have glandular linings that secrete nutrients for nourishment and maintenance of the sperm. The ejaculatory duct is derived from an invagination of the epidermal cells during development and, as a result, has a cuticular lining. The terminal portion of the ejaculatory duct may be sclerotized to form the intromittent organ, the aedeagus. The remainder of the male reproductive system is derived from embryonic mesoderm, except for the germ cells, or spermatogonia, which descend from the primordial pole cells very early during embryogenesis. The aedeagus can be quite pronounced or de minimis. The base of the aedeagus may be the partially sclerotized phallotheca, also called the phallosoma or theca. In some species the phallotheca contains a space, called the endosoma (internal holding pouch), into which the tip end of the aedeagus may be withdrawn (retracted). The vas deferens is sometimes drawn into (folded into) the phallotheca together with a seminal vesicle.
Blattodea
Cockroaches are most common in tropical and subtropical climates. Some species are in close association with human dwellings and widely found around garbage or in the kitchen. Cockroaches are generally omnivorous with the exception of the wood-eating species such as Cryptocercus; these roaches are incapable of digesting cellulose themselves, but have symbiotic relationships with various protozoans and bacteria that digest the cellulose, allowing them to extract the nutrients. The similarity of these symbionts in the genus Cryptocercus to those in termites are such that it has been suggested that they are more closely related to termites than to other cockroaches, and current research strongly supports this hypothesis of relationships. All species studied so far carry the obligate mutualistic endosymbiont bacterium Blattabacterium, with the exception of Nocticola australiensis, an Australian cave dwelling species without eyes, pigment or wings, and which recent genetic studies indicates are very primitive cockroaches.
Cockroaches, like all insects, breathe through a system of tubes called tracheae. The tracheae of insects are attached to the spiracles, excluding the head. Thus cockroaches, like all insects, are not dependent on the mouth and windpipe to breathe. The valves open when the CO2 level in the insect rises to a high level; then the CO2 diffuses out of the tracheae to the outside and fresh O2 diffuses in. Unlike in vertebrates that depend on blood for transporting O2 and CO2, the tracheal system brings the air directly to cells, the tracheal tubes branching continually like a tree until their finest divisions, tracheoles, are associated with each cell, allowing gaseous oxygen to dissolve in the cytoplasm lying across the fine cuticle lining of the tracheole. CO2 diffuses out of the cell into the tracheole. While cockroaches do not have lungs and thus do not actively breathe in the vertebrate lung manner, in some very large species the body musculature may contract rhythmically to forcibly move air out and in the spiracles; this may be considered a form of breathing.
Coleoptera
The digestive system of beetles is primarily based on plants, which they for the most part feed upon, with mostly the anterior midgut performing digestion. However, in predatory species (e.g., Carabidae) most digestion occurs in the crop by means of midgut enzymes. In Elateridae species, the predatory larvae defecate enzymes on their prey, with digestion being extraorally. The alimentary canal basically comprises a short narrow pharynx, a widened expansion, the crop and a poorly developed gizzard. After there is a midgut, that varies in dimensions between species, with a large amount of cecum, with a hingut, with varying lengths. There are typically four to six Malpighian tubules.
The nervous system in beetles contains all the types found in insects, varying between different species. With three thoracic and seven or eight abdominal ganglia can be distinguished to that in which all the thoracic and abdominal ganglia are fused to form a composite structure. Oxygen is obtained via a tracheal system. Air enters a series of tubes along the body through openings called spiracles, and is then taken into increasingly finer fibers. Pumping movements of the body force the air through the system. Some species of diving beetles (Dytiscidae) carry a bubble of air with them whenever they dive beneath the water surface. This bubble may be held under the elytra or it may be trapped against the body using specialized hairs. The bubble usually covers one or more spiracles so the insect can breathe air from the bubble while submerged. An air bubble provides an insect with only a short-term supply of oxygen, but thanks to its unique physical properties, oxygen will diffuse into the bubble and displacing the nitrogen, called passive diffusion, however the volume of the bubble eventually diminishes and the beetle will have to return to the surface.
Like other insect species, beetles have hemolymph instead of blood. The open circulatory system of the beetle is driven by a tube-like heart attached to the top inside of the thorax.
Different glands specialize for different pheromones produced for finding mates. Pheromones from species of Rutelinea are produced from epithelial cells lining the inner surface of the apical abdominal segments or amino acid based pheromones of Melolonthinae from eversible glands on the abdominal apex. Other species produce different types of pheromones. Dermestids produce esters, and species of Elateridae produce fatty-acid-derived aldehydes and acetates. For means of finding a mate also, fireflies (Lampyridae) utilized modified fat body cells with transparent surfaces backed with reflective uric acid crystals to biosynthetically produce light, or bioluminescence. The light produce is highly efficient, as it is produced by oxidation of luciferin by the enzymes luciferase in the presence of ATP (adenosine triphospate) and oxygen, producing oxyluciferin, carbon dioxide, and light.
A notable number of species have developed special glands that produce chemicals for deterring predators (see Defense and predation). The Ground beetle's (of Carabidae) defensive glands, located at the posterior, produce a variety of hydrocarbons, aldehydes, phenols, quinones, esters, and acids released from an opening at the end of the abdomen. While African carabid beetles (e.g., Anthia some of which used to comprise the genus Thermophilum) employ the same chemicals as ants: formic acid. While Bombardier beetles have well developed, like other carabid beetles, pygidial glands that empty from the lateral edges of the intersegment membranes between the seventh and eighth abdominal segments. The gland is made of two containing chambers. The first holds hydroquinones and hydrogen peroxide, with the second holding just hydrogen peroxide plus catalases. These chemicals mix and result in an explosive ejection, forming temperatures of around 100 C, with the breakdown of hydroquinone to H2 + O2 + quinone, with the O2 propelling the excretion.
Tympanal organs are hearing organs. Such an organ is generally a membrane (tympanum) stretched across a frame backed by an air sac and associated sensory neurons. In the order Coleoptera, tympanal organs have been described in at least two families. Several species of the genus Cicindela in the family Cicindelidae have ears on the dorsal surface of the first abdominal segment beneath the wing; two tribes in the family Dynastinae (Scarabaeidae) have ears just beneath the pronotal shield or neck membrane. The ears of both families are to ultrasonic frequencies, with strong evidence that they function to detect the presence of bats via their ultrasonic echolocation. Even though beetles constitute a large order and live in a variety of niches, examples of hearing is surprisingly lacking in species, though it is likely that most are just undiscovered.
Dermaptera
The neuroendocrine system is typical of insects. There is a brain, a subesophageal ganglion, three thoracic ganglia, and six abdominal ganglia. Strong neuron connections connect the neurohemal corpora cardiaca to the brain and frontal ganglion, where the closely related median corpus allatum produces juvenile hormone III in close proximity to the neurohemal dorsal arota. The digestive system of earwigs is like all other insects, consisting of a fore-, mid-, and hindgut, but earwigs lack gastric caecae which are specialized for digestion in many species of insect. Long, slender (extratory) malpighian tubules can be found between the junction of the mid- and hind gut.
The reproductive system of females consist of paired ovaries, lateral oviducts, spermatheca, and a genital chamber. The lateral ducts are where the eggs leave the body, while the spermatheca is where sperm is stored. Unlike other insects, the gonopore, or genital opening is behind the seventh abdominal segment. The ovaries are primitive in that they are polytrophic (the nurse cells and oocytes alternate along the length of the ovariole). In some species these long ovarioles branch off the lateral duct, while in others, short ovarioles appear around the duct.
Diptera
The genitalia of female flies are rotated to a varying degree from the position found in other insects. In some flies this is a temporary rotation during mating, but in others it is a permanent torsion of the organs that occurs during the pupal stage. This torsion may lead to the anus being located below the genitals, or, in the case of 360° torsion, to the sperm duct being wrapped around the gut, despite the external organs being in their usual position. When flies mate, the male initially flies on top of the female, facing in the same direction, but then turns round to face in the opposite direction. This forces the male to lie on its back in order for its genitalia to remain engaged with those of the female, or the torsion of the male genitals allows the male to mate while remaining upright. This leads to flies having more reproduction abilities than most insects and at a much quicker rate. Flies come in great populations due ir ability to mate effectively and in a short period of time especially during the mating season.
The female lays her eggs as close to the food source as possible, and development is very rapid, allowing the larva to consume as much food as possible in a short period of time before transforming into the adult. The eggs hatch immediately after being laid, or the flies are ovoviviparous, with the larva hatching inside the mother. Larval flies, or maggots, have no true legs, and little demarcation between the thorax and abdomen; in the more derived species, the head is not clearly distinguishable from the rest of the body. Maggots are limbless, or else have small prolegs. The eyes and antennae are reduced or absent, and the abdomen also lacks appendages such as cerci. This lack of features is an adaptation to a food-rich environment, such as within rotting organic matter, or as an endoparasite. The pupae take various forms, and in some cases develop inside a silk cocoon. After emerging from the pupa, the adult fly rarely lives more than a few days, and serves mainly to reproduce and to disperse in search of new food sources.
Lepidoptera
In reproductive system of butterflies and moths, the male genitalia are complex and unclear. In females there are three types of genitalia based on the relating taxa: monotrysian, exoporian, and dytresian. In the monotrysian type there is an opening on the fused segments of the sterna 9 and 10, which act as insemination and oviposition. In the exoporian type (in Hepaloidae and Mnesarchaeoidea) there are two separate places for insemination and oviposition, both occurring on the same sterna as the monotrysian type, 9/10. In most species the genitalia are flanked by two soft lobes, although they may be specialized and sclerotized in some species for ovipositing in area such as crevices and inside plant tissue. Hormones and the glands that produce them run the development of butterflies and moths as they go through their life cycle, called the endocrine system. The first insect hormone PTTH (Prothoracicotropic hormone) operates the species life cycle and diapause (see the relates section). This hormone is produced by corpora allata and corpora cardiaca, where it is also stored. Some glands are specialized to perform certain task such as producing silk or producing saliva in the palpi. While the corpora cardiaca produce PTTH, the corpora allata also produces jeuvanile hormones, and the prothorocic glands produce moulting hormones.
In the digestive system, the anterior region of the foregut has been modified to form a pharyngial sucking pump as they need it for the food they eat, which are for the most part liquids. An esophagus follows and leads to the posterior of the pharynx and in some species forms a form of crop. The midgut is short and straight, with the hindgut being longer and coiled. Ancestors of lepidopteran species, stemming from Hymenoptera, had midgut ceca, although this is lost in current butterflies and moths. Instead, all the digestive enzymes other than initial digestion, are immobilized at the surface of the midgut cells. In larvae, long-necked and stalked goblet cells are found in the anterior and posterior midgut regions, respectively. In insects, the goblet cells excrete positive potassium ions, which are absorbed from leaves ingested by the larvae. Most butterflies and moths display the usual digestive cycle, however species that have a different diet require adaptations to meet these new demands.
In the circulatory system, hemolymph, or insect blood, is used to circulate heat in a form of thermoregulation, where muscles contraction produces heat, which is transferred to the rest of the body when conditions are unfavorable. In lepidopteran species, hemolymph is circulated through the veins in the wings by some form of pulsating organ, either by the heart or by the intake of air into the trachea. Air is taken in through spiracles along the sides of the abdomen and thorax supplying the trachea with oxygen as it goes through the lepidopteran's respiratory system. There are three different tracheae supplying oxygen diffusing oxygen throughout the species body: The dorsal, ventral, and visceral. The dorsal tracheae supply oxygen to the dorsal musculature and vessels, while the ventral tracheae supply the ventral musculature and nerve cord, and the visceral tracheae supply the guts, fat bodies, and gonads.