
Scientific name Protozoa | Subclass Coccidia | |
 | ||
Lower classifications Rhizopoda, Sarcodina, parasitic protozoans, Sarcomastigophora, Mycetozoa Similar Bacteria , Protozoan infection , Eukaryote | ||
Protozoa classification
In 21st-century systems of biological classification, the Protozoa are defined as a diverse group of unicellular eukaryotic organisms. Historically, protozoa were defined as single-celled animals or organisms with animal-like behaviors, such as motility and predation. The group was regarded as the zoological counterpart to the "protophyta", which were considered to be plant-like, as they are capable of photosynthesis.
Contents
- Protozoa classification
- Protozoa characteristics of protozoa 01 02
- History and terminology
- Characteristics
- Motility and feeding
- Pellicle
- Life cycle
- Classification
- Ecological role
- In humans
- In other animals
- References
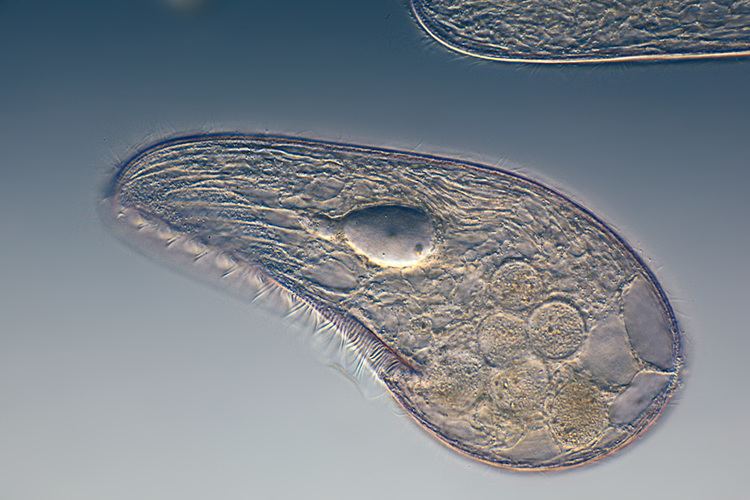
The terms protozoa and protozoans are now mostly used informally to designate single-celled, non-photosynthetic protists, such as the ciliates, amoebae and flagellates.
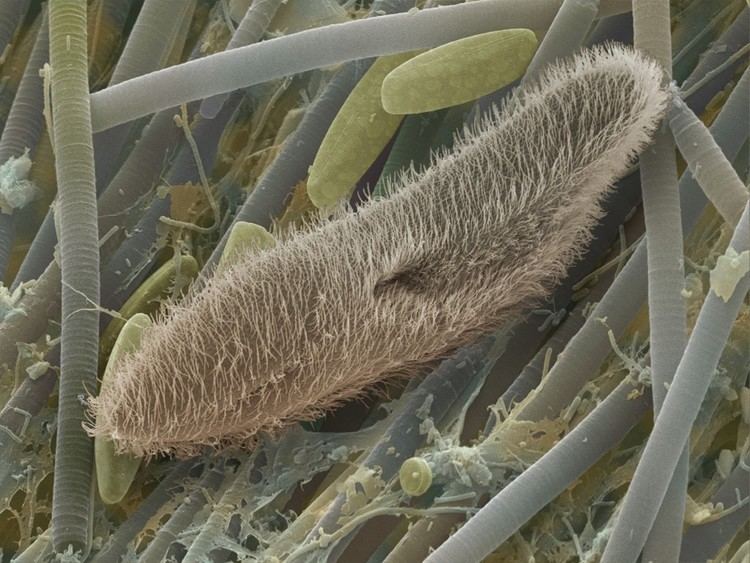
The term Protozoa was introduced in 1818 for a taxonomic class, but in later classification schemes the group was elevated to higher ranks, including phylum, subkingdom and kingdom. In several classification systems proposed by Thomas Cavalier-Smith and his collaborators since 1981, Protozoa is ranked as a kingdom. The seven-kingdom scheme proposed by Ruggiero et al. in 2015, places eight phyla under Protozoa: Euglenozoa, Amoebozoa, Metamonada, Choanozoa, Loukozoa, Percolozoa, Microsporidia and Sulcozoa. This kingdom does not form a clade, but an evolutionary grade or paraphyletic group, from which the fungi and animals are specifically excluded.
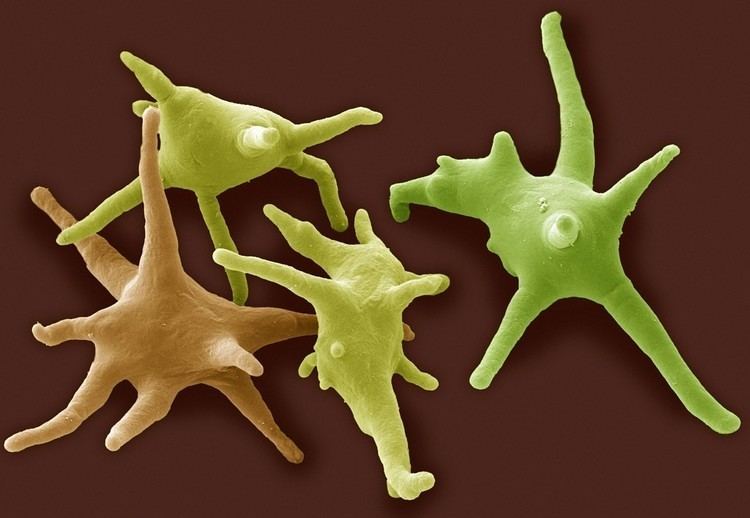
The use of Protozoa as a formal taxon has been discouraged by some recent researchers, mainly because the term, which is formed from the Greek protos "first" + zoia, plural of zoion, "animal", implies kinship with animals (metazoa) and promotes a separation of "animal-like" from "plant-like" organisms, neither of which they encourage. Modern ultrastructural, biochemical, and genetic techniques have shown that protozoa, as traditionally defined, belong to widely divergent lineages, and can no longer be regarded as "primitive animals." For this reason, the terms "protists," "Protista" or "Protoctista" are sometimes preferred for the high-level classification of eukaryotic microbes. In 2005, members of the Society of Protozoologists voted to change the name of that organization to the International Society of Protistologists.
Protozoa characteristics of protozoa 01 02
History and terminology
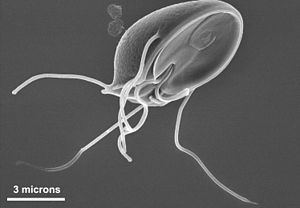
The word "protozoa" (singular protozoon or protozoan) was coined in 1818 by zoologist Georg August Goldfuss, as the Greek equivalent of the German Urthiere, meaning "primitive, or original animals" (ur- ‘proto-’ + Thier ‘animal’). Goldfuss erected Protozoa as a class containing what he believed to be the simplest animals. Originally, the group included not only microbes, but also some "lower" multicellular animals, such as rotifers, corals, sponges, jellyfish, bryozoa and polychaete worms.
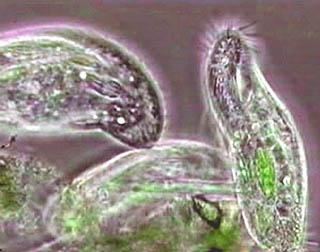
In 1848, in light of advancements in cell theory pioneered by Theodore Schwann and Matthias Schleiden, the anatomist and zoologist C.T. von Siebold proposed that the bodies of microbial organisms such as ciliates and amoebae were made up of single cells, similar to those from which the multicellular tissues of plants and animals were constructed. Von Siebold redefined Protozoa to include only such unicellular forms, to the exclusion of all metazoa. At the same time, he raised the group to the level of a phylum containing two broad classes of microbes: Infusoria (mostly ciliates and flagellated algae), and Rhizopoda (amoeboid organisms). The definition of Protozoa as a phylum or sub-kingdom made up of "unicellular animals" was adopted by the zoologist Otto Bütschli—celebrated at his centenary as the "architect of protozoology"—and the term came into wide use.
As a phylum under Animalia, the Protozoa were firmly rooted in the old "two-kingdom" classification of life, according to which all living beings were classified as either animals or plants. As long as this scheme remained dominant, the protozoa were understood to be animals and studied in departments of Zoology, while photosynthetic microbes and microscopic fungi—the so-called Protophyta—were assigned to the Plants, and studied in departments of Botany.
Criticism of this system began in the latter half of the 19th century, with the realization that many organisms met the criteria for inclusion among both plants and animals. For example, the algae Euglena and Dinobryon have chloroplasts for photosynthesis, but can also feed on organic matter and are motile. In 1860, John Hogg argued against the use of "protozoa", on the grounds that "naturalists are divided in opinion—and probably some will ever continue so—whether many of these organisms, or living beings, are animals or plants." As an alternative, he proposed a new kingdom called Primigenum, consisting of both the protozoa and unicellular algae (protophyta), which he combined together under the name "Protoctista". In Hoggs's conception, the animal and plant kingdoms were likened to two great "pyramids" blending at their bases in the Kingdom Primigenum.
Six years later, Ernst Haeckel also proposed a third kingdom of life, which he named Protista. At first, Haeckel included a few multicellular organisms in this kingdom, but in later work he restricted the Protista to single-celled organisms, or simple colonies whose individual cells are not differentiated into different kinds of tissues.
Despite these proposals, Protozoa emerged as the preferred taxonomic placement for heterotrophic microbes such as amoebae and ciliates, and remained so for more than a century. In the course of the 20th century, however, the old "two kingdom" system began to weaken, with the growing awareness that fungi did not belong among the plants, and that most of the unicellular protozoa were no more closely related to the animals than they were to the plants. By mid-century, some biologists, such as Herbert Copeland, Robert H. Whittaker and Lynn Margulis, advocated the revival of Haeckel's Protista or Hogg's Protoctista as a kingdom-level eukaryotic group, alongside Plants, Animals and Fungi. A variety of multi-kingdom systems were proposed, and Kingdoms Protista and Protoctista became well established in biology texts and curricula.
While many taxonomists have abandoned Protozoa as a high-level group, Thomas Cavalier-Smith has retained it as a kingdom in the various classifications he has proposed. As of 2015, Cavalier-Smith's Protozoa excludes several major groups of organisms traditionally placed among the protozoa, including the ciliates, dinoflagellates and foraminifera (all members of the SAR supergroup). In its current form, his kingdom Protozoa is a paraphyletic group which includes a common ancestor and most of its descendents, but excludes two important clades that branch within it: the animals and fungi.
Characteristics
Protozoa, as traditionally defined, are mainly microscopic organisms, ranging in size from 10 to 52 micrometers. Some, however, are significantly larger. Among the largest are the deep-sea–dwelling xenophyophores, single-celled foraminifera whose shells can reach 20 cm in diameter. Free-living forms are restricted to moist environments, such as soils, mosses and aquatic habitats, although many form resting cysts which enable them to survive drying. Many protozoan species are symbionts, some are parasites, and some are predators of bacteria, algae and other protists.
Motility and feeding
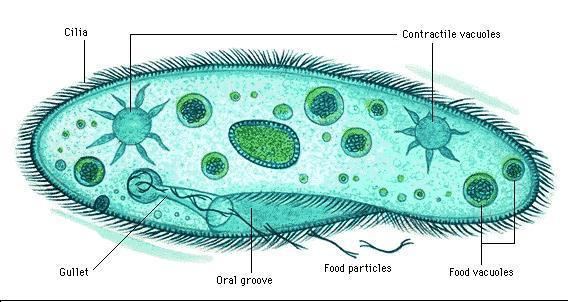
Organisms traditionally classified as protozoa are abundant in aqueous environments and soil, occupying a range of trophic levels. The group includes flagellates (which move with the help of whip-like structures called flagella), ciliates (which move by using hair-like structures called cilia) and amoebae (which move by the use of foot-like structures called pseudopodia). Some protozoa are sessile, and do not move at all.
Protozoa may take in food by osmotrophy, absorbing nutrients through their cell membranes; or they may feed by phagocytosis, either by engulfing particles of food with pseudopodia (as amoebae do), or taking in food through a mouth-like aperture called a cytostome. All protozoa digest their food in stomach-like compartments called vacuoles.
Pellicle
The pellicle is a thin layer supporting the cell membrane in various protozoa, such as ciliates, protecting them and allowing them to retain their shape, especially during locomotion, allowing the organism to be more hydrodynamic. The pellicle varies from flexible and elastic to rigid. Although somewhat stiff, the pellicle is also flexible and allows the protist to fit into tighter spaces. In ciliates and Apicomplexa, it is formed from closely packed vesicles called alveoli. In euglenids, it is formed from protein strips arranged spirally along the length of the body. Familiar examples of protists with a pellicle are the euglenoids and the ciliate Paramecium. In some protozoa, the pellicle hosts epibiotic bacteria that adhere to the surface by their fimbriae (attachment pili).
Life cycle
Some protozoa have life phases alternating between proliferative stages (e.g., trophozoites) and dormant cysts. As cysts, protozoa can survive harsh conditions, such as exposure to extreme temperatures or harmful chemicals, or long periods without access to nutrients, water, or oxygen for periods of time. Being a cyst enables Parasitic species to survive outside of a host, and allows their transmission from one host to another. When protozoa are in the form of trophozoites (Greek tropho = to nourish), they actively feed. The conversion of a trophozoite to cyst form is known as encystation, while the process of transforming back into a trophozoite is known as excystation. Protozoa reproduce asexually by binary fission or multiple fission. Many protozoan species exchange genetic material by sexual means (typically, through conjugation); however, sexuality is generally decoupled from the process of reproduction, and does not immediately result in increased population.
Although meiotic sex is widespread among present day eukaryotes, it has, until recently, been unclear whether or not eukaryotes were sexual early in their evolution. Due to recent advances in gene detection and other techniques, evidence has been found for some form of meiotic sex in an increasing number of protozoans of ancient lineage that diverged early in eukaryotic evolution. (See eukaryote reproduction.) Thus, such findings suggest that meiotic sex arose early in eukaryotic evolution. Examples of protozoan meiotic sexuality are described in the articles Amoebozoa, Giardia lamblia, Leishmania, Plasmodium falciparum biology, Paramecium, Toxoplasma gondii, Trichomonas vaginalis and Trypanosoma brucei.
Classification
The classification of protozoa has been and remains a problematic area of taxonomy. Where they are available, DNA sequences are used as the basis for classification; however, for the majority of described protozoa, such material is not available. Protozoa have been and still are organized mostly on the basis of their morphology, means of locomotion, and for the parasitic species their hosts.
As a phylum the Protozoa were, historically, divided into four subphyla reflecting the means of locomotion:
These systems are no longer considered to be valid.
Ecological role
As components of the micro- and meiofauna, protozoa are an important food source for microinvertebrates. Thus, the ecological role of protozoa in the transfer of bacterial and algal production to successive trophic levels is important. As predators, they prey upon unicellular or filamentous algae, bacteria, and microfungi. Protozoan species include both herbivores and consumers in the decomposer link of the food chain. They also control bacteria populations and biomass to some extent. On average, protozoa eat ~ 100 to 1,000 bacteria per hour. Protozoa can stimulate decomposition of organic matter, digest cellulose in the rumen of cows and termite guts, and can play a role in nutrient mobilization.
In humans
A number of protozoan pathogens are human parasites, causing diseases such as malaria (by Plasmodium), amoebiasis, giardiasis, toxoplasmosis, cryptosporidiosis, trichomoniasis, Chagas disease, leishmaniasis, African trypanosomiasis (sleeping sickness), amoebic dysentery, acanthamoeba keratitis, and primary amoebic meningoencephalitis (naegleriasis).
In other animals
The protozoan Ophryocystis elektroscirrha is a parasite of butterfly larvae, passed from female to caterpillar. Severely infected individuals are weak, unable to expand their wings, or unable to eclose, and have shortened lifespans, but parasite levels vary in populations. Infection creates a culling effect, whereby infected migrating animals are less likely to complete the migration. This results in populations with lower parasite loads at the end of the migration. This is not the case in laboratory or commercial rearing, where after a few generations, all individuals can be infected.