
Scientific name Fabaceae | ||
 | ||
Lower classifications | ||
Class 11 12 fabaceae family
The Fabaceae, Leguminosae or Papilionaceae, commonly known as the legume, pea, or bean family, are a large and economically important family of flowering plants. It includes trees, shrubs, and perennial or annual herbaceous plants, which are easily recognized by their fruit (legume) and their compound, stipulated leaves. The family is widely distributed, and is the third-largest land plant family in terms of number of species, behind only the Orchidaceae and Asteraceae, with about 751 genera and some 19,000 known species . The five largest of the genera are Astragalus (over 3,000 species), Acacia (over 1000 species), Indigofera (around 700 species), Crotalaria (around 700 species) and Mimosa (around 500 species), which constitute about a quarter of all legume species. The ca. 19,000 known legume species amount to about 7% of flowering plant species. Fabaceae is the most common family found in tropical rainforests and in dry forests in the Americas and Africa.
Contents
- Class 11 12 fabaceae family
- Taxonomy and diversity of fabaceae
- Etymology
- Description
- Growth habit
- Leaves
- Roots
- Flowers
- Fruit
- Physiology and biochemistry
- Distribution and habitat
- Biological nitrogen fixation
- Evolution
- Phylogeny
- Taxonomy
- Genera
- Economic and cultural importance
- Food and forage
- Natural gums
- Dyes
- Ornamentals
- Emblematic Leguminosae
- References

Recent molecular and morphological evidence supports the fact that the Fabaceae is a single monophyletic family. This point of view has been supported not only by the degree of interrelation shown by different groups within the family compared with that found among the Leguminosae and their closest relations, but also by all the recent phylogenetic studies based on DNA sequences. These studies confirm that the Fabaceae are a monophyletic group that is closely related to the Polygalaceae, Surianaceae and Quillajaceae families and that they belong to the order Fabales.

Along with the cereals, some fruits and tropical roots a number of Leguminosae have been a staple human food for millennia and their use is closely related to human evolution.

A number are important agricultural and food plants, including Glycine max (soybean), Phaseolus (beans), Pisum sativum (pea), Cicer arietinum (chickpeas), Medicago sativa (alfalfa), Arachis hypogaea (peanut), Lathyrus odoratus (sweet pea), Ceratonia siliqua (carob), and Glycyrrhiza glabra (liquorice). A number of species are also weedy pests in different parts of the world, including: Cytisus scoparius (broom), Robinia pseudoacacia (black locust), Ulex europaeus (gorse), Pueraria lobata (Kudzu), and a number of Lupinus species.
Taxonomy and diversity of fabaceae
Etymology

The name 'Fabaceae' comes from the defunct genus Faba, now included in Vicia. The term "faba" comes from Latin, and appears to simply mean "bean". Leguminosae is an older name still considered valid, and refers to the fruit of these plants, which are called legumes.
Description
Fabaceae range in habit from giant trees (like Koompassia excelsa) to small annual herbs, with the majority being herbaceous perennials. Plants have indeterminate inflorescences, which are sometimes reduced to a single flower. The flowers have a short hypanthium and a single carpel with a short gynophore, and after fertilization produce fruits that are legumes.
Growth habit
The Leguminosae have a wide variety of growth forms including trees, shrubs or herbaceous plants or even vines or lianas. The herbaceous plants can be annuals, biennials or perennials, without basal or terminal leaf aggregations. They are upright plants, epiphytes or vines. The latter support themselves by means of shoots that twist around a support or through cauline or foliar tendrils. Plants can be heliophytes, mesophytes or xerophytes.
Leaves
The leaves are usually alternate and compound. Most often they are even- or odd-pinnately compound (e.g. Caragana and Robinia respectively), often trifoliate (e.g. Trifolium, Medicago) and rarely palmately compound (e.g. Lupinus), in the Mimosoideae and the Caesalpinioideae commonly bipinnate (e.g. Acacia, Mimosa). They always have stipules, which can be leaf-like (e.g. Pisum), thorn-like (e.g. Robinia) or be rather inconspicuous. Leaf margins are entire or, occasionally, serrate. Both the leaves and the leaflets often have wrinkled pulvini to permit nastic movements. In some species, leaflets have evolved into tendrils (e.g. Vicia).
Many species have leaves with structures that attract ants that protect the plant from herbivore insects (a form of mutualism). Extrafloral nectaries are common among the Mimosoideae and the Caesalpinioideae, and are also found in some Faboideae (e.g. Vicia sativa). In some Acacia, the modified hollow stipules are inhabited by ants and are known as domatia.
Roots
Many Fabaceae host bacteria in their roots within structures called root nodules. These bacteria, known as rhizobia, have the ability to take nitrogen gas (N2) out of the air and convert it to a form of nitrogen that is usable to the host plant ( NO3− or NH3 ). This process is called nitrogen fixation. The legume, acting as a host, and rhizobia, acting as a provider of usable nitrate, form a symbiotic relationship.
Flowers
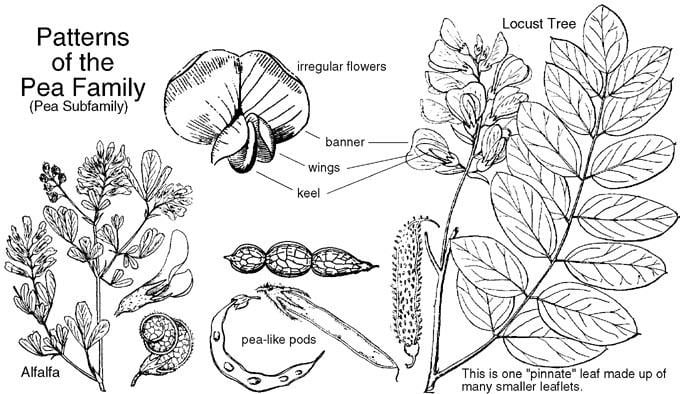
The flowers often have five generally fused sepals and five free petals. They are generally hermaphrodite, and have a short hypanthium, usually cup shaped. There are normally ten stamens and one elongated superior ovary, with a curved style. They are usually arranged in indeterminate inflorescences. Fabaceae are typically entomophilous plants (i.e. they are pollinated by insects), and the flowers are usually showy to attract pollinators.
In the Caesalpinioideae, the flowers are often zygomorphic, as in Cercis, or nearly symmetrical with five equal petals in Bauhinia. The upper petal is the innermost one, unlike in the Faboideae. Some species, like some in the genus Senna, have asymmetric flowers, with one of the lower petals larger than the opposing one, and the style bent to one side. The calyx, corolla, or stamens can be showy in this group.
In the Mimosoideae, the flowers are actinomorphic and arranged in globose inflorescences. The petals are small and the stamens, which can be more than just 10, have long, coloured filaments, which are the showiest part of the flower. All of the flowers in an inflorescence open at once.
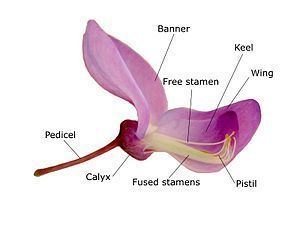
In the Faboideae, the flowers are zygomorphic, and have a specialized structure. The upper petal, called the banner, is large and envelops the rest of the petals in bud, often reflexing when the flower blooms. The two adjacent petals, the wings, surround the two bottom petals. The two bottom petals are fused together at the apex (remaining free at the base), forming a boat-like structure called the keel. The stamens are always ten in number, and their filaments can be fused in various configurations, often in a group of nine stamens plus one separate stamen. Various genes in the CYCLOIDEA (CYC)/DICHOTOMA (DICH) family are expressed in the upper (also called dorsal or adaxial) petal; in some species, such as Cadia, these genes are expressed throughout the flower, producing a radially symmetrical flower.
Fruit
The ovary most typically develops into a legume. A legume is a simple dry fruit that usually dehisces (opens along a seam) on two sides. A common name for this type of fruit is a "pod", although that can also be applied to a few other fruit types. A few species have evolved samarae, loments, follicles, indehiscent legumes, achenes, drupes, and berries from the basic legume fruit.
Physiology and biochemistry
The Leguminosae are rarely cyanogenic, however, where they are, the cyanogenic compounds are derived from tyrosine, phenylalanine or leucine. They frequently contain alkaloids. Proanthocyanidins can be present either as cyanidin or delphinidine or both at the same time. Flavonoids such as kaempferol, quercitin and myricetin are often present. Ellagic acid has never been found in any of the genera or species analysed. Sugars are transported within the plants in the form of sucrose. C3 photosynthesis has been found in a wide variety of genera. The family has also evolved a unique chemistry. Pterocarpans are a class of molecules (derivatives of isoflavonoids) found only in the Fabaceae.
Distribution and habitat
The Fabaceae have an essentially worldwide distribution, being found everywhere except Antarctica and the high arctic. The trees are often found in tropical regions, while the herbaceous plants and shrubs are predominant outside the tropics.
Biological nitrogen fixation
Biological nitrogen fixation (BNF, performed by the organisms called diazotrophs) is a very old process that probably originated in the Archean eon when the primitive atmosphere lacked oxygen. It is only carried out by Euryarchaeota and just 6 of the more than 50 phyla of bacteria. Some of these lineages co-evolved together with the flowering plants establishing the molecular basis of a mutually beneficial symbiotic relationship. BNF is carried out in nodules that are mainly located in the root cortex, although they are occasionally located in the stem as in Sesbania rostrata. The spermatophytes that co-evolved with actinorhizal diazotrophs (Frankia) or with rhizobia to establish their symbiotic relationship belong to 11 families contained within the Rosidae clade (as established by the gene molecular phylogeny of rbcL, a gene coding for part of the RuBisCO enzyme in the chloroplast). This grouping indicates that the predisposition for forming nodules probably only arose once in flowering plants and that it can be considered as an ancestral characteristic that has been conserved or lost in certain lineages. However, such a wide distribution of families and genera within this lineage indicates that nodulation had multiple origins. Of the 10 families within the Rosidae, 8 have nodules formed by actinomyces (Betulaceae, Casuarinaceae, Coriariaceae, Datiscaceae, Elaeagnaceae, Myricaceae, Rhamnaceae and Rosaceae), and the two remaining families, Ulmaceae and Fabaceae have nodules formed by rhizobia.
The rhizobia and their hosts must be able to recognize each other for nodule formation to commence. Rhizobia are specific to particular host species although a rhizobia species may often infect more than one host species. This means that one plant species may be infected by more than one species of bacteria. For example, nodules in Acacia senegal can contain seven species of rhizobia belonging to three different genera. The most distinctive characteristics that allow rhizobia to be distinguished apart are the rapidity of their growth and the type of root nodule that they form with their host. Root nodules can be classified as being either indeterminate, cylindrical and often branched, and determinate, spherical with prominent lenticels. Indeterminate nodules are characteristic of legumes from temperate climates, while determinate nodules are commonly found in species from tropical or subtropical climates.
Nodule formation is common throughout the leguminosae, it is found in the majority of its members that only form an association with rhizobia, which in turn form an exclusive symbiosis with the leguminosae (with the exception of Parasponia, the only genus of the 18 Ulmaceae genera that is capable of forming nodules). Nodule formation is present in all the leguminosae sub-families, although it is less common in the Caesalpinioideae. All types of nodule formation are present in the sub-family Papilionoideae: indeterminate (with the meristem retained), determinate (without meristem) and the type included in Aeschynomene. The latter two are thought to be the most modern and specialised type of nodule as they are only present in some lines of the Papilionoideae sub-family. Even though nodule formation is common in the two monophyletic subfamilies Papilionoideae and Mimosoideae they also contain species that do not form nodules. The presence or absence of nodule-forming species within the three sub-families indicates that nodule formation has arisen several times during the evolution of the leguminosae and that this ability has been lost in some lineages. For example, within the genus Acacia, a member of the Mimosoideae, A. pentagona does not form nodules, while other species of the same genus readily form nodules, as is the case for Acacia senegal, which forms both rapidly and slow growing rhizobial nodules.
Evolution
The order Fabales contains around 7.3% of eudicot species and the greatest part of this Diversity is contained in just one of the four families that order contains: Fabaceae. This clade also includes the Polygalaceae, Surianaceae and Quillajaceae families and its origins date back 94 to 89 million years, although it started its diversification some 79 to 74 million years ago. In fact, the Fabaceae have diversified during the early Tertiary to become a ubiquitous part of the modern earth’S biota, along with many other families belonging to the flowering plants.
The Fabaceae have an abundant and diverse fossil record, especially for the Tertiary period. Fossils of flowers, fruit, leaves, wood and pollen from this period have been found in numerous locations. The earliest fossils that can be definitively assigned to the Fabaceae appeared in the late Palaeocene (approximately 56 million years ago). Representatives of the 3 sub-families traditionally recognised as being members of the Fabaceae – Cesalpinioideae, Papilionoideae and Mimosoideae — as well as members of the large clades within these sub-families – such as the genistoides – have been found in periods a little later, starting between 55 and 50 million years ago. In fact, a wide variety of taxa representing the main lineages in the Fabaceae have been found in the fossil record dating from the middle to the late Eocene, suggesting that the majority of the modern Fabaceae groups were already present and that a broad diversification occurred during this period. Therefore, the Fabaceae started their diversification approximately 60 million years ago and the most important clades separated some 50 million years ago. The age of the main Cesalpinioideae clades have been estimated as between 56 and 34 million years and the basal group of the Mimosoideae as 44 ± 2.6 million years. The division between Mimosoideae and Faboideae is dated as occurring between 59 and 34 million years ago and the basal group of the Faboideae as 58.6 ± 0.2 million years ago. It has been possible to date the divergence of some of the groups within the Faboideae, even though diversification within each genus was relatively recent. For instance, Astragalus separated from the Oxytropis some 16 to 12 million years ago. In addition, the separation of the aneuploid species of Neoastragalus started 4 million years ago. Inga, another genus of the Papilionoideae with approximately 350 species, seems to have diverged in the last 2 million years.
It has been suggested, based on fossil and phylogenetic evidence, that legumes originally evolved in arid and/or semi-arid regions along the Tethys seaway during the Palaeogene Period. However, others contend that Africa (or even the Americas) cannot yet be ruled out as the origin of the family.
The current hypothesis about the evolution of the genes needed for nodulation is that they were recruited from other pathways after a polyploidy event. Several different pathways have been implicated as donating duplicated genes to the pathways need for nodulation. The main donors to the pathway were the genes associated with the arbuscular mycorrhiza symbiosis genes, the pollen tube formation genes and the haemoglobin genes. One of the main genes shown to be shared between the arbuscular mycorrhiza pathway and the nodulation pathway is SYMRK and it is involved in the plant-bacterial recognition. The pollen tube growth is similar to the infection thread development in that infection threads grow in a polar manner that is similar to a pollen tubes polar growth towards the ovules. Both pathways include the same type of enzymes, pectin-degrading cell wall enzymes. The enzymes needed to reduce nitrogen, nitrogenases, require a substantial input of ATP but at the same time are sensitive to free oxygen. To meet the requirements of this paradoxical situation, the plants express a type of haemoglobin called leghaemoglobin that is believed to be recruited after a duplication event. These three genetic pathways are believed to be part of a gene duplication event then recruited to work in nodulation.
Phylogeny
The phylogeny of the legumes has been the object of many studies by research groups from around the world. These studies have used morphology, DNA data (the chloroplast intron trnL, the chloroplast genes rbcL and matK, or the ribosomal spacers ITS) and cladistic analysis in order to investigate the relationships between the family’s different lineages. Fabaceae is consistently recovered as monophyletic. The studies further confirmed that the traditional subfamilies Mimosoideae and Papilionoideae were each monophyletic but both were nested within the paraphyletic subfamily Caesalpinioideae. All the different approaches yielded similar results regarding the relationships between the family's main clades. Following extensive discussion in the legume phylogenetics community, the Legume Phylogeny Working Group reclassified Fabaceae into six subfamilies, which necessitated the segregation of four new subfamilies from Caesalpinioideae and merging Caesapinioideae sensu stricto with the former subfamily Mimosoideae.
Taxonomy
The Fabaceae are placed in the order Fabales according to most taxonomic systems, including the APG III system. The family now includes six subfamilies:
Genera
The 730 genera included in this family can be viewed on the following three pages:
Economic and cultural importance
Legumes are economically and culturally important plants due to their extraordinary diversity and abundance, the wide variety of edible vegetables they represent and due to the variety of uses they can be put to: in horticulture and agriculture, as a food, for the compounds they contain that have medicinal uses and for the oil and fats they contain that have a variety of uses.
Food and forage
The history of legumes is tied in closely with that of human civilization, appearing early in Asia, the Americas (the common bean, several varieties) and Europe (broad beans) by 6,000 BCE, where they became a staple, essential as a source of protein.
Their ability to fix atmospheric nitrogen reduces fertilizer costs for farmers and gardeners who grow legumes, and means that legumes can be used in a crop rotation to replenish soil that has been depleted of nitrogen. Legume seeds and foliage have a comparatively higher protein content than non-legume materials, due to the additional nitrogen that legumes receive through the process. Some legume species perform hydraulic lift, which makes them ideal for intercropping.
Farmed legumes can belong to numerous classes, including forage, grain, blooms, pharmaceutical/industrial, fallow/green manure and timber species, with most commercially farmed species filling two or more roles simultaneously.
There are of two broad types of forage legumes. Some, like alfalfa, clover, vetch, and Arachis, are sown in pasture and grazed by livestock. Other forage legumes such as Leucaena or Albizia are woody shrub or tree species that are either broken down by livestock or regularly cut by humans to provide stock feed.
Grain legumes are cultivated for their seeds, and are also called pulses. The seeds are used for human and animal consumption or for the production of oils for industrial uses. Grain legumes include both herbaceous plants like beans, lentils, lupins, peas and peanuts. and trees such as carob, mesquite and tamarind.
Bloom legume species include species such as lupin, which are farmed commercially for their blooms as well as being popular in gardens worldwide. Laburnum, Robinia, Gleditsia, Acacia, Mimosa, and Delonix are ornamental trees and shrubs.
Industrial farmed legumes include Indigofera, cultivated for the production of indigo, Acacia, for gum arabic, and Derris, for the insecticide action of rotenone, a compound it produces.
Fallow or green manure legume species are cultivated to be tilled back into the soil to exploit the high nitrogen levels found in most legumes. Numerous legumes are farmed for this purpose, including Leucaena, Cyamopsis and Sesbania.
Various legume species are farmed for timber production worldwide, including numerous Acacia species, Dalbergia species, and Castanospermum australe.
Melliferous plants offer nectar to bees and other insects to encourage them to carry pollen from the flowers of one plant to others thereby ensuring pollination.A number of legume species are good nectar providers such as alfalfa, white clover, sweet clover and various Prosopis species. Many plants in the Fabaceae family are an important source of pollen for the bumblebee species Bombus hortorum. This bee species is especially fond of one species in particular; Trifolium pratense, also known as red clover, is a popular food source in the diet of Bombus hortorum.
Natural gums
Natural gums are vegetable exudates that are released as the result of damage to the plant such as that resulting from the attack of an insect or a natural or artificial cut. These exudates contain heterogeneous polysaccharides formed of different sugars and usually containing uronic acids. They form viscous colloidal solutions. There are different species that produce gums. The most important of these species belong to the leguminosae. They are widely used in the pharmaceutical, cosmetic, food and textile sectors. They also have interesting therapeutic properties; for example gum arabic is antitussive and anti-inflammatory. The most well known gums are tragacanth (Astragalus gummifer), gum arabic (Acacia senegal) and guar gum (Cyamopsis tetragonoloba).
Dyes
The species used to produce dyes include the following: Logwood Haematoxylon campechianum; a large spiny tree that can grow up to 15 m tall. Its cork is thin and soft and its wood is hard. The heartwood is used to produce dyes that are red and purple. The histological stain called haematoxylin is produced from this species. Brazilwood tree (Caesalpinia echinata) is similar to the previous tree but smaller and with red or purple flowers. The wood is also used to produce a red or purple dye. The Madras thorn (Pithecallobium dulce) is another spiny tree native to Latin America, it grows up to 4 m high and has yellow or green flowers that grow in florets. Its fruit is reddish and is used to produce a yellow dye. Indigo dye is extracted from the True indigo plant Indigofera tinctoria that is native to Asia. In Central and South America dyes are produced from two species related to this species, indigo from Indigofera suffruticosa and Natal indigo from Indigofera arrecta.yellow dye is extracted from Butea monosperma commonly called as flame of the forest.
Ornamentals
Legumes have been used as ornamental plants throughout the world for many centuries. Their vast diversity of heights, shapes, foliage and flower colour means that this family is commonly used in the design and planting of everything from small gardens to large parks. The following is a list of the main ornamental legume species, listed by sub-family.