
Scientific name Eukaryota | ||
 | ||
Lower classifications Plant, Animal, protozoa, Ciliate, Dinosaur Similar Animal, Ciliate, Dinosaur | ||
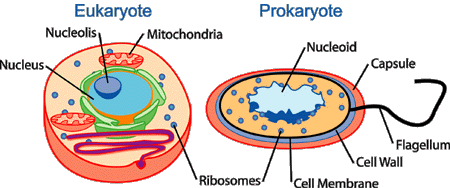
Prokaryotes and eukaryotes
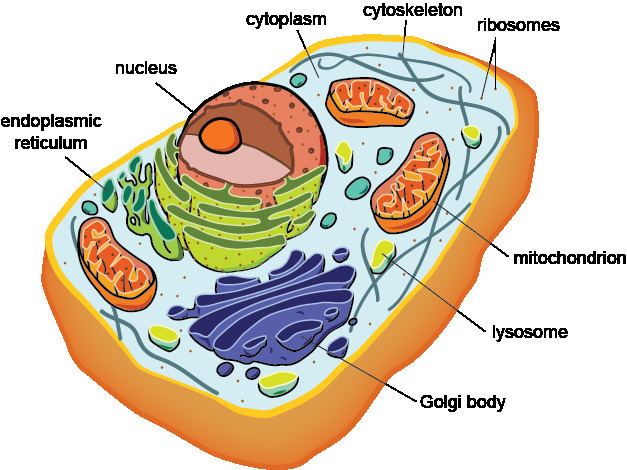
A eukaryote (/juːˈkæri.oʊt/ or /juːˈkæriət/ yoo-KARR-ee-oht or yoo-KARR-ee-ət) is any organism whose cells contain a nucleus and other organelles enclosed within membranes. Eukaryotes belong to the taxon Eukarya or Eukaryota. The defining feature that sets eukaryotic cells apart from prokaryotic cells (Bacteria and Archaea) is that they have membrane-bound organelles, especially the nucleus, which contains the genetic material and is enclosed by the nuclear envelope. The presence of a nucleus gives eukaryotes their name, which comes from the Greek εὖ (eu, "well" or "true") and κάρυον (karyon, "nut" or "kernel"). Eukaryotic cells also contain other membrane-bound organelles such as mitochondria and the Golgi apparatus. In addition, plants and algae contain chloroplasts. Eukaryotic organisms may be unicellular or multicellular. Only eukaryotes form multicellular organisms consisting of many kinds of tissue made up of different cell types.
Contents
- Prokaryotes and eukaryotes
- Prokaryote vs eukaryote
- Cell features
- Internal membrane
- Mitochondria and plastids
- Cytoskeletal structures
- Cell wall
- Differences among eukaryotic cells
- Animal cell
- Plant cell
- Fungal cell
- Other eukaryotic cells
- Reproduction
- Classification
- Phylogeny
- Five supergroups
- Cavalier Smiths tree
- Alternative views
- Fossils
- Relationship to Archaea
- Endomembrane system and mitochondria
- Hypotheses for the origin of eukaryotes
- Autogenous models
- Chimeric models
- References

Eukaryotes can reproduce both asexually through mitosis and sexually through meiosis and gamete fusion. In mitosis, one cell divides to produce two genetically identical cells. In meiosis, DNA replication is followed by two rounds of cell division to produce four daughter cells each with half the number of chromosomes as the original parent cell (haploid cells). These act as sex cells (gametes – each gamete has just one complement of chromosomes, each a unique mix of the corresponding pair of parental chromosomes) resulting from genetic recombination during meiosis.
The domain Eukaryota appears to be monophyletic, and so makes up one of the three domains of life. The two other domains, bacteria and Archaea, are prokaryotes and have none of the above features. Eukaryotes represent a tiny minority of all living things. However, due to their much larger size, eukaryotes' collective worldwide biomass is estimated at about equal to that of prokaryotes. Eukaryotes first developed approximately 1.6–2.1 billion years ago (during the Proterozoic eon).
Prokaryote vs eukaryote
Cell features
Eukaryotic cells are typically much larger than those of prokaryotes. They have a variety of internal membrane-bound structures, called organelles, and a cytoskeleton composed of microtubules, microfilaments, and intermediate filaments, which play an important role in defining the cell's organization and shape. Eukaryotic DNA is divided into several linear bundles called chromosomes, which are separated by a microtubular spindle during nuclear division.
Internal membrane
Eukaryote cells include a variety of membrane-bound structures, collectively referred to as the endomembrane system. Simple compartments, called vesicles or vacuoles, can form by budding off other membranes. Many cells ingest food and other materials through a process of endocytosis, where the outer membrane invaginates and then pinches off to form a vesicle. It is probable that most other membrane-bound organelles are ultimately derived from such vesicles. Alternatively some products produced by the cell can leave in a vesicle through exocytosis.
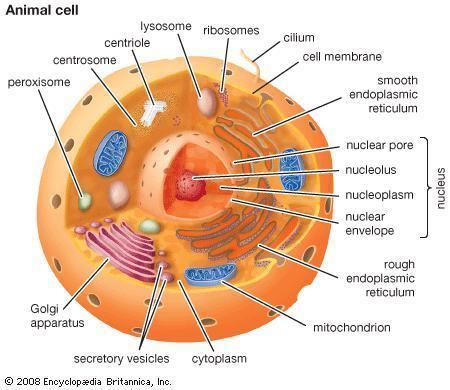
The nucleus is surrounded by a double membrane (commonly referred to as a nuclear membrane or nuclear envelope), with pores that allow material to move in and out. Various tube- and sheet-like extensions of the nuclear membrane form what is called the endoplasmic reticulum or ER, which is involved in protein transport and maturation. It includes the rough ER where ribosomes are attached to synthesize proteins, which enter the interior space or lumen. Subsequently, they generally enter vesicles, which bud off from the smooth ER. In most eukaryotes, these protein-carrying vesicles are released and further modified in stacks of flattened vesicles, called Golgi bodies or dictyosomes.
Vesicles may be specialized for various purposes. For instance, lysosomes contain digestive enzymes that break down the contents of food vacuoles, and Peroxisomes are used to break down peroxide, which is toxic otherwise. Many protozoa have contractile vacuoles, which collect and expel excess water, and extrusomes, which expel material used to deflect predators or capture prey. In higher plants, most of a cell's volume is taken up by a central vacuole, which primarily maintains its osmotic pressure.
Mitochondria and plastids
Mitochondria are organelles found in nearly all eukaryotes that provide energy to the cell by converting ingested sugars into ATP. They are surrounded by two membranes (each a phospholipid bi-layer), the inner of which is folded into invaginations called cristae, where aerobic respiration takes place. Mitochondria contain their own DNA. They are now generally held to have developed from endosymbiotic prokaryotes, probably proteobacteria. Protozoa and microbes that lack mitochondria, such as the amoebozoan Pelomyxa and metamonads such as Giardia and Trichomonas, have usually been found to contain mitochondrion-derived organelles, such as hydrogenosomes and mitosomes, and thus probably lost the mitochondria secondarily.
In 2016, Monocercomonoides, a metamonad flagellate which resides in the intestines of the chinchilla, has been found to lack mitochondria entirely. Monocercomonoides obtains its energy by enzymatic action on nutrients absorbed from the environment. It has also acquired, by lateral Gene transfer, a cytosolic sulphur mobilisation system which provides the clusters of iron and sulfur required for protein synthesis. The normal mitochondrial iron-sulphur cluster pathway is considered to have been lost secondarily.
Plants and various groups of algae also have plastids. plastids have their own DNA and are developed from endosymbionts, in this case cyanobacteria. They usually take the form of chloroplasts, which like cyanobacteria contain chlorophyll and produce organic compounds (such as glucose) through photosynthesis. Others are involved in storing food. Although plastids probably had a single origin, not all plastid-containing groups are closely related. Instead, some eukaryotes have obtained them from others through secondary endosymbiosis or ingestion.
Endosymbiotic origins have also been proposed for the nucleus, for which see below, and for eukaryotic flagella, supposed to have developed from spirochaetes. This is not generally accepted, both from a lack of cytological evidence and difficulty in reconciling this with cellular reproduction.
Cytoskeletal structures
Many eukaryotes have long slender motile cytoplasmic projections, called flagella, or similar structures called cilia. Flagella and cilia are sometimes referred to as undulipodia, and are variously involved in movement, feeding, and sensation. They are composed mainly of tubulin. These are entirely distinct from prokaryotic flagellae. They are supported by a bundle of microtubules arising from a basal body, also called a kinetosome or centriole, characteristically arranged as nine doublets surrounding two singlets. Flagella also may have hairs, or mastigonemes, and scales connecting membranes and internal rods. Their interior is continuous with the cell's cytoplasm.
Microfilamental structures composed of actin and actin binding proteins, e.g., α-actinin, fimbrin, filamin are Present in submembraneous cortical layers and bundles, as well. Motor proteins of microtubules, e.g., dynein or kinesin and actin, e.g., myosins provide dynamic character of the network.
centrioles are often present even in cells and groups that do not have flagella, but Conifers and flowering plants have neither. They generally occur in groups of one or two, called kinetids, that give rise to various microtubular roots. These form a primary component of the cytoskeletal structure, and are often assembled over the course of several cell divisions, with one flagellum retained from the parent and the other derived from it. Centrioles may also be associated in the formation of a spindle during nuclear division.
The significance of cytoskeletal structures is underlined in the determination of shape of the cells, as well as their being essential components of migratory responses like chemotaxis and chemokinesis. Some protists have various other microtubule-supported organelles. These include the radiolaria and heliozoa, which produce axopodia used in flotation or to capture prey, and the haptophytes, which have a peculiar flagellum-like organelle called the haptonema.
Cell wall
The cells of plants, fungi, and most chromalveolates have a cell wall, a layer outside the cell membrane, providing the cell with structural support, protection, and a filtering mechanism. The cell wall also prevents over-expansion when water enters the cell.
The major polysaccharides making up the primary cell wall of Land plants are cellulose, hemicellulose, and pectin. The cellulose microfibrils are linked via hemicellulosic tethers to form the cellulose-hemicellulose network, which is embedded in the pectin matrix. The most common hemicellulose in the primary cell wall is xyloglucan.
Differences among eukaryotic cells
There are many different types of eukaryotic cells, though Animals and plants are the most familiar eukaryotes, and thus provide an excellent starting point for understanding eukaryotic structure. Fungi and many protists have some substantial differences, however.
Animal cell
All animals consist of eukaryotic cells. Animal cells are distinct from those of other eukaryotes, most notably plants, as they lack cell walls and chloroplasts and have smaller vacuoles. Due to the lack of a cell wall, animal cells can adopt a variety of shapes. A phagocytic cell can even engulf other structures.
There are many other types of cell. For instance, there are approximately 210 distinct cell types in the adult human body.
Plant cell
Plant cells are quite different from the cells of the other eukaryotic organisms. Their distinctive features are:
Fungal cell
Fungal cells are most similar to animal cells, with the following exceptions:
Other eukaryotic cells
Eukaryotes are a very diverse group, and their cell structures are equally diverse. Many have cell walls; many do not. Many have chloroplasts, derived from primary, secondary, or even tertiary endosymbiosis; and many do not. Some groups have unique structures, such as the cyanelles of the glaucophytes, the haptonema of the haptophytes, or the ejectisomes of the cryptomonads. Other structures, such as pseudopods, are found in various eukaryote groups in different forms, such as the lobose amoebozoans or the reticulose foraminiferans.
Reproduction
Cell division generally takes place asexually by mitosis, a process that allows each daughter nucleus to receive one copy of each chromosome. In most eukaryotes, there is also a process of sexual reproduction, typically involving an alternation between haploid generations, wherein only one copy of each chromosome is present, and diploid generations, wherein two copies of each chromosome are present, occurring through meiosis. There is considerable variation in this pattern.
Eukaryotes have a smaller surface area to volume ratio than prokaryotes, and thus have lower metabolic rates and longer generation times. In some multicellular organisms, cells specialized for metabolism will have enlarged surface areas, such as intestinal vili.
The evolution of sexual reproduction may be a primordial and fundamental characteristic of eukaryotes. Based on a phylogenetic analysis, Dacks and Roger proposed that facultative sex was present in the common ancestor of all eukaryotes. A core set of genes that function in meiosis is present in both Trichomonas vaginalis and Giardia intestinalis, two organisms previously thought to be asexual. Since these two species are descendants of lineages that diverged early from the eukaryotic evolutionary tree, it was inferred that core meiotic genes, and hence sex, were likely present in a common ancestor of all eukaryotes. Other studies on eukaryotic species once thought to be asexual have revealed evidence for a sexual cycle. For instance, parasitic protozoa of the genus Leishmania have recently been shown to have a sexual cycle. Also, evidence now indicates that amoeba, that were previously regarded as asexual, are anciently sexual and that the majority of present-day asexual groups likely arose recently and independently.
Classification
In antiquity, the two clades of animals and plants were recognized. They were given the taxonomic rank of Kingdom by Linnaeus. Though he included the fungi with plants with some reservations, it was later realized that they are quite distinct and warrant a separate kingdom, the composition of which was not entirely clear until the 1980s. The various single-cell eukaryotes were originally placed with plants or animals when they became known. In 1830, the German biologist Georg A. Goldfuss coined the word protozoa to refer to organisms such as ciliates, and this group was expanded until it encompassed all single-celled eukaryotes, and given their own kingdom, the Protista, by Ernst Haeckel in 1866. The eukaryotes thus came to be composed of four kingdoms:
The protists were understood to be "primitive forms", and thus an evolutionary grade, united by their primitive unicellular nature. The disentanglement of the deep splits in the tree of life only really started with DNA sequencing, leading to a system of domains rather than kingdoms as top level rank being put forward by Carl Woese, uniting all the eukaryote kingdoms under the eukaryote domain. At the same time, work on the protist tree intensified, and is still actively going on today. Several alternative classifications have been forwarded, though there is no consensus in the field.
A classification produced in 2005 for the International Society of Protistologists, which reflected the consensus of the time, divided the eukaryotes into six supposedly monophyletic 'supergroups'. However, in the same year (2005), doubts were expressed as to whether some of these supergroups were monophyletic, particularly the Chromalveolata, and a review in 2006 noted the lack of evidence for several of the supposed six supergroups. A revised classification in 2012 recognizes five supergroups.
There are also smaller groups of eukaryotes whose position is uncertain or seems to fall outside the major groups. In particular, Haptophyta, Cryptophyta, Centrohelida, Telonemia, Picozoa, Apusomonadida, Ancyromonadida, Breviatea, and the genus Collodictyon. Overall, it seems that, although progress has been made, there are still very significant uncertainties in the evolutionary history and classification of eukaryotes. As Roger & Simpson said in 2009 "with the current pace of change in our understanding of the eukaryote tree of life, we should proceed with caution."
In an article published in Nature Microbiology in April 2016 the authors, "reinforced once again that the life we see around us – plants, animals, humans and other so-called eukaryotes – represent a tiny percentage of the world's biodiversity." They classified eukaryote "based on the inheritance of their information systems as opposed to lipid or other cellular structures." Jillian F. Banfield of the University of California, Berkeley and fellow scientists used a super computer to generate a diagram of a new tree of life based on DNA from 3000 species including 2,072 known species and 1,011 newly reported microbial organisms, whose DNA they had gathered from diverse environments. As the capacity to sequence DNA became easier, Banfield and team were able to do metagenomic sequencing—"sequencing whole communities of organisms at once and picking out the individual groups based on their genes alone."
Phylogeny
The rRNA trees constructed during the 1980s and 1990s left most eukaryotes in an unresolved "crown" group (not technically a true crown), which was usually divided by the form of the mitochondrial cristae; see crown eukaryotes. The few groups that lack mitochondria branched separately, and so the absence was believed to be primitive; but this is now considered an artifact of long-branch attraction, and they are known to have lost them secondarily.
As of 2011, there is widespread agreement that the Rhizaria belong with the Stramenopiles and the Alveolata, in a clade dubbed the SAR supergroup, so that Rhizaria is not one of the main eukaryote groups; also that the Amoebozoa and Opisthokonta are each monophyletic and form a clade, often called the unikonts. Beyond this, there does not appear to be a consensus.
It has been estimated that there may be 75 distinct lineages of eukaryotes. Most of these lineages are protists.
The known eukaryote genome sizes vary from 8.2 megabases (Mb) in Babesia bovis to 112,000–220,050 Mb in the dinoflagellate Prorocentrum micans, suggesting that the genome of the ancestral eukaryote has undergone considerable variation during its evolution. The last common ancestor of all eukaryotes is believed to have been a phagotrophic protist with a nucleus, at least one centriole and cilium, facultatively aerobic mitochondria, sex (meiosis and syngamy), a dormant cyst with a cell wall of chitin and/or cellulose and peroxisomes. Later endosymbiosis led to the spread of plastids in some lineages.
Five supergroups
A global tree of eukaryotes from a consensus of phylogenetic evidence (in particular, phylogenomics), rare genomic signatures, and morphological characteristics is presented in Adl et al. 2012 and Burki 2014.
In some analyses, the Hacrobia group (Haptophyta + Cryptophyta) is placed next to Archaeplastida, but in other ones it is nested inside the Archaeplastida. However, several recent studies have concluded that Haptophyta and Cryptophyta do not form a monophyletic group. The former could be a sister group to the SAR group, the latter cluster with the Archaeplastida (plants in the broad sense). As of February 2012, it remains unclear whether the Hacrobia forms a monophyletic group.
The division of the eukaryotes into two primary clades, bikonts (Archaeplastida + SAR + Excavata) and unikonts (Amoebozoa + Opisthokonta), derived from an ancestral biflagellar organism and an ancestral uniflagellar organism, respectively, had been suggested earlier. A 2012 study produced a somewhat similar division, although noting that the terms "unikonts" and "bikonts" were not used in the original sense.
Cavalier-Smith's tree
Cavalier-Smith 2010, 2013, and 2014 places the eukaryotic tree's root between Excavata (with ventral feeding groove supported by a microtubular root) and the grooveless Euglenozoa:
Alternative views
Other analyses place the SAR supergroup within an expanded Chromalveolata, although they differ on the placement of the resulting five groups. Rogozin et al. in 2009 produced the tree shown below, where the primary division is between the Archaeplastida and all other eukaryotes.
A paper published in 2009, which re-examined the data used in some of the analyses presented above as well as performing new ones, strongly suggested that the Archaeplastida are polyphyletic. The phylogeny finally proposed in the paper is shown below.
There are also the cladistic analyses of Diana Lipscomb based on classical data which have Red algae as basal and Archeoplastida as paraphyletic. In the survey by Parfrey et al. it is recovered in only 42.6% of the molecular analyses that include it, that is, 26 out of 61. It is not recovered in Goloboff et al.'s combined analysis, and is mostly weakly supported in other molecular analyses. Laura Parfrey et al. point out that Archeoplastida support comes primarily from phylogenomic analyses and these may be picking up misleading endosymbiotic gene transfer signal of genes independently transferred from the plastid to the host nucleus in the 3 archeoplastid clades. And Stiller and Harrell emphasize that the group can be explained by "short-branch exclusion" and "subtle and easily overlooked biases can dominate the overall results of molecular phylogenetic analyses of ancient eukaryotic relationships. Sources of potential phylogenetic artifact should be investigated routinely, not just when obvious 'long-branch attraction' is encountered."
Red algae as basal is also supported by molecular evidence. Cyanidioschyzon, a red alga, is considered basal (sister group to the rest of eukaryotes) by Nagashima et al. and Seckbach. It has the most primitive chloroplast, only 1 mitochondrion, no vacuoles, no trienoic acids, and the smallest eukaryotic genome at 8 Mbp.
Rhizaria are only moderately supported (65.5% of the studies, i.e., 19 of 29), statistical support for them is inconsistent in multigene genealogies with larger taxon sampling and the group is ambiguously supported in Goloboff et al.
Fossils
The origin of the eukaryotic cell is considered a milestone in the evolution of life, since eukaryotes include all complex cells and almost all multicellular organisms. The timing of this series of events is hard to determine; Knoll (2006) suggests they developed approximately 1.6–2.1 billion years ago. Some acritarchs are known from at least 1.65 billion years ago, and the possible alga Grypania has been found as far back as 2.1 billion years ago. The Geosiphon-like fossil fungus Diskagma has been found in paleosols 2.2 billion years old
Organized living structures have been found in the black shales of the Palaeoproterozoic Francevillian B Formation in Gabon, dated at 2.1 billion years old. Eukaryotic life could have evolved at that time. Fossils that are clearly related to modern groups start appearing an estimated 1.2 billion years ago, in the form of a red alga, though recent work suggests the existence of fossilized filamentous algae in the Vindhya basin dating back perhaps to 1.6 to 1.7 billion years ago.
Biomarkers suggest that at least stem eukaryotes arose even earlier. The presence of steranes in Australian shales indicates that eukaryotes were present in these rocks dated at 2.7 billion years old.
Relationship to Archaea
Eukaryotes are more closely related to Archaea than Bacteria, at least in terms of nuclear DNA and genetic machinery, and one controversial idea is to place them with Archaea in the clade Neomura. However, in other respects, such as membrane composition, eukaryotes are similar to Bacteria. Three main explanations for this have been proposed:
The chronocyte hypothesis for the origin of the eukaryotic cell postulates that a primitive eukaryotic cell was formed by the endosymbiosis of both archaea and bacteria by a third type of cell, termed a chronocyte.
A proposed cladogram for the positioning of the Eukaryotes is, with the Eukaryote root in the excavates and the Eukaryotes as sister of the Heimdallarchaeota,
Endomembrane system and mitochondria
The origins of the endomembrane system and mitochondria are also unclear. The phagotrophic hypothesis proposes that eukaryotic-type membranes lacking a cell wall originated first, with the development of endocytosis, whereas mitochondria were acquired by ingestion as endosymbionts. The syntrophic hypothesis proposes that the proto-eukaryote relied on the proto-mitochondrion for food, and so ultimately grew to surround it. Here the membranes originated after the engulfment of the mitochondrion, in part thanks to mitochondrial genes (the hydrogen hypothesis is one particular version).
In a study using genomes to construct supertrees, Pisani et al. (2007) suggest that, along with evidence that there was never a mitochondrion-less eukaryote, eukaryotes evolved from a syntrophy between an archaea closely related to Thermoplasmatales and an α-proteobacterium, likely a symbiosis driven by sulfur or hydrogen. The mitochondrion and its genome is a remnant of the α-proteobacterial endosymbiont.
Hypotheses for the origin of eukaryotes
Different hypotheses have been proposed as to how eukaryotic cells came into existence. These hypotheses can be classified into two distinct classes – autogenous models and chimeric models.
Autogenous models
Autogenous models propose that a proto-eukaryotic cell containing a nucleus existed first, and later acquired mitochondria. According to this model, a large prokaryote developed invaginations in its plasma membrane in order to obtain enough surface area to service its cytoplasmic volume. As the invaginations differentiated in function, some became separate compartments—giving rise to the endomembrane system, including the endoplasmic reticulum, Golgi apparatus, nuclear membrane, and single membrane structures such as lysosomes. Mitochondria are proposed to come from the endosymbiosis of an aerobic proteobacterium, and it's assumed that all the eukaryotic lineages that did not acquire mitochondria became extinct. Chloroplasts came about from another endosymbiotic event involving cyanobacteria. Since all eukaryotes have mitochondria, but not all have chloroplasts, mitochondria are thought to have come first. This is the serial endosymbiosis theory.
Some models propose that the origins of double layered organelles, such as mitochondria and chloroplasts, in the proto-eukaryotic cell is due to the compartmentalization of DNA vesicles that were formed from the secondary invaginations or more detailed infoldings of cellular membrane.
Chimeric models
Chimeric models claim that two prokaryotic cells existed initially – an archaeon and a bacterium. These cells underwent a merging process, either by a physical fusion or by endosymbiosis, thereby leading to the formation of a eukaryotic cell. Within these chimeric models, some studies further claim that mitochondria originated from a bacterial ancestor while others emphasize the role of endosymbiotic processes behind the origin of mitochondria.
Based on the process of mutualistic symbiosis, the hypotheses can be categorized as – the serial endosymbiotic theory (SET), the hydrogen hypothesis (mostly a process of symbiosis where hydrogen transfer takes place among different species), and the syntrophy hypothesis.
According to serial endosymbiotic theory (championed by Dr. Lynn Margulis), a union between a motile anaerobic bacterium (like Spirochaeta) and a thermoacidophilic crenarchaeon (like Thermoplasma which is sulfidogenic in nature) gave rise to the present day eukaryotes. This union established a motile organism capable of living in the already existing acidic and sulfurous waters. Oxygen is known to cause toxicity to organisms that lack the required metabolic machinery. Thus, the archaeon provided the bacterium with a highly beneficial reduced environment (sulfur and sulfate were reduced to sulfide). In microaerophilic conditions, oxygen was reduced to water thereby creating a mutual benefit platform. The bacterium on the other hand, contributed the necessary fermentation products and electron acceptors along with its motility feature to the archaeon thereby gaining a swimming motility for the organism. From a consortium of bacterial and archaeal DNA originated the nuclear genome of eukaryotic cells. Spirochetes gave rise to the motile features of eukaryotic cells. Endosymbiotic unifications of the ancestors of alpha-proteobacteria and cyanobacteria, led to the origin of mitochondria and plastids respectively. For example, Thiodendron has been known to have originated via an ectosymbiotic process based on a similar syntrophy of sulfur existing between the two types of bacteria – Desulphobacter and Spirochaeta. However, such an association based on motile symbiosis have never been observed practically. Also there is no evidence of archaeans and spirochetes adapting to intense acid-based environments.
In the hydrogen hypothesis, the symbiotic linkage of an anaerobic and autotrophic methanogenic archaeon (host) with an alpha-proteobacterium (the symbiont) gave rise to the eukaryotes. The host utilized hydrogen (H2) and carbon dioxide (CO2) to produce methane while the symbiont, capable of aerobic respiration, expelled H2 and CO2 as byproducts of anaerobic fermentation process. The host's methanogenic environment worked as a sink for H2, which resulted in heightened bacterial fermentation. Endosymbiotic gene transfer (EGT) acted as a catalyst for the host to acquire the symbionts' carbohydrate metabolism and turn heterotrophic in nature. Subsequently, the host's methane forming capability was lost. Thus, the origins of the heterotrophic organelle (symbiont) are identical to the origins of the eukaryotic lineage. In this hypothesis, the presence of H2 represents the selective force that forged eukaryotes out of prokaryotes.
The syntrophy hypothesis was developed in contrast to the hydrogen hypothesis and proposes the existence of two symbiotic events. According to this theory, eukaryogenesis (i.e. origin of eukaryotic cells) occurred based on metabolic symbiosis (syntrophy) between a methanogenic archaeon and a delta-proteobacterium. This syntrophic symbiosis was initially facilitated by H2 transfer between different species under anaerobic environments. In earlier stages, an alpha-proteobacterium became a member of this integration, and later developed into the mitochondrion. Gene transfer from a delta-proteobacterium to an archaeon led to the methanogenic archaeon developing into a nucleus. The archaeon constituted the genetic apparatus while the delta-proteobacterium contributed towards the cytoplasmic features. This theory incorporates two selective forces that were needed to be considered during the time of nucleus evolution – (a) presence of metabolic partitioning in order to avoid the harmful effects of the co-existence of anabolic and catabolic cellular pathways, and (b) prevention of abnormal biosynthesis of proteins that occur due to a vast spread of introns in the archaeal genes after acquiring the mitochondrion and the loss of methanogenesis.
Thus, the origin of eukaryotes by endosymbiotic processes has been broadly recognized and accepted so far. Mitochondria and plastids have been known to originate from a bacterial ancestor during parallel adaptation to anaerobiosis. However, there still remains a greater need in assessing the question of how much eukaryotic complexity is being originated via an implementation of these symbiogenetic theories.