
Domain Bacteria Family Enterobacteriaceae Higher classification Yersinia | Kingdom Eubacteria Order Enterobacteriales Genus Yersinia Rank Species | |
 | ||
Similar | ||
Yersinia pestis the plague microbiology lectures
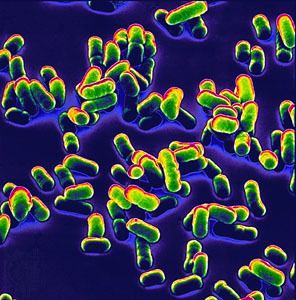
Yersinia pestis (formerly Pasteurella pestis) is a gram-negative, rod-shaped coccobacillus, a facultative anaerobic organism that can infect humans via the Oriental rat flea. It causes the deadly disease called bubonic plague (or "the plague" colloquially). Human Y. pestis infection takes three main forms: pneumonic, septicemic, and bubonic plagues. All three forms were responsible for a number of high-mortality epidemics throughout human history, including: the sixth century's Plague of Justinian; the Black Death, which accounted for the death of at least one-third of the European population between 1347 and 1353; and the 19th century's Third Pandemic. These plagues probably originated in China and were transmitted west via trade routes.
Contents
- Yersinia pestis the plague microbiology lectures
- General characteristics
- Genome
- Small non coding RNA
- See also
- Pathogenesis and immunity
- In reservoir hosts
- Vector
- In humans and other susceptible hosts
- Immunity
- Symptoms and disease progression
- Clinical determination
- Treatment
- Prevention
- Plague of Justinian
- Role in Black Death
- Use in biological warfare
- Proposed use in biological warfare
- Recent events
- References
Y. pestis was discovered in 1894 by Alexandre Yersin, a Swiss/French physician and bacteriologist from the Pasteur Institute, during an epidemic of the plague in Hong Kong. Yersin was a member of the Pasteur school of thought. Kitasato Shibasaburō, a German-trained Japanese bacteriologist who practised Koch's methodology, was also engaged at the time in finding the causative agent of the plague. However, Yersin actually linked plague with Y. pestis. Named Pasteurella pestis in the past, the organism was renamed Yersinia pestis in 1944.

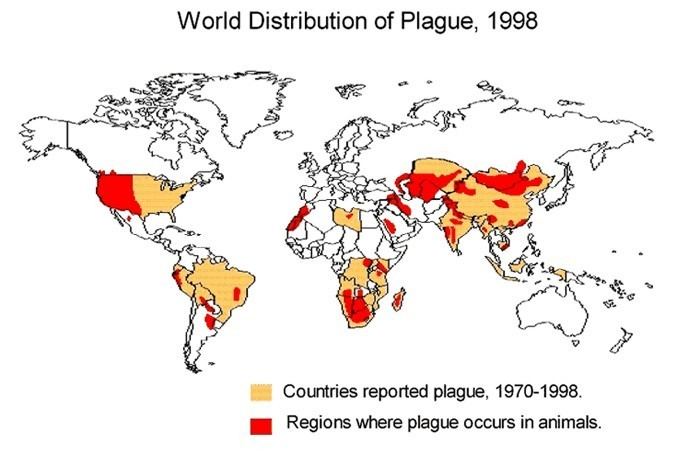
Every year, thousands of cases of the plague are still reported to the World Health Organization, although, with proper treatment, the prognosis for victims is now much better. A five- to six-fold increase in cases occurred in Asia during the time of the Vietnam War, possibly due to the disruption of ecosystems and closer proximity between people and animals. The plague also has a detrimental effect on non-human mammals. In the United States, mammals such as the black-tailed prairie dog and the endangered black-footed ferret are under threat.
General characteristics

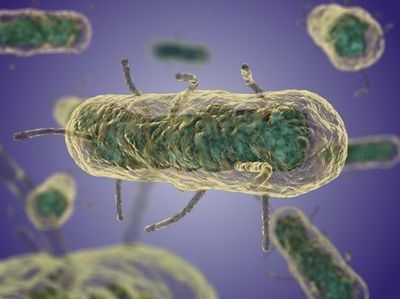
Y. pestis is a non-motile, stick-shaped, facultative anaerobic bacterium with bipolar staining (giving it a safety pin appearance) that produces an anti-phagocytic slime layer. Similar to other Yersinia species, it tests negative for urease, lactose fermentation, and indole. The closest relative is the gastrointestinal pathogen Yersinia pseudotuberculosis, and more distantly Yersinia enterocolitica.
Genome
The complete genomic sequence is available for two of the three subspecies of Y. pestis: strain KIM (of biovar Y. p. medievalis), and strain CO92 (of biovar Y. p. orientalis, obtained from a clinical isolate in the United States). As of 2006, the genomic sequence of a strain of biovar Antiqua has been recently completed. Similar to the other pathogenic strains, there are signs of loss of function mutations. The chromosome of strain KIM is 4,600,755 base pairs long; the chromosome of strain CO92 is 4,653,728 base pairs long. Like Y. pseudotuberculosis and Y. enterocolitica, Y. pestis is host to the plasmid pCD1. In addition, it also hosts two other Plasmids, pPCP1 (also called pPla or pPst) and pMT1 (also called pFra) that are not carried by the other Yersinia species. pFra codes for a phospholipase D that is important for the ability of Y. pestis to be transmitted by fleas. pPla codes for a protease, Pla, that activates plasmin in human hosts and is a very important virulence factor for pneumonic plague. Together, these plasmids, and a pathogenicity island called HPI, encode several proteins that cause the Pathogenesis, for which Y. pestis is famous. Among other things, these virulence factors are required for bacterial adhesion and injection of proteins into the host Cell, invasion of bacteria in the host cell (via a type-III secretion system), and acquisition and binding of iron harvested from red blood cells (by siderophores). Y. pestis is thought to be descendant from Y. pseudotuberculosis, differing only in the presence of specific virulence plasmids.

A comprehensive and comparative proteomics analysis of Y. pestis strain KIM was performed in 2006. The analysis focused on the transition to a growth condition mimicking growth in host cells.
Small non-coding RNA
Numerous bacterial small non-coding RNAs have been identified to play regulatory functions. Some can regulate the virulence genes. 63 novel putative sRNA were identified through deep sequencing of the Y. pestis sRNA-ome. Among them was Yersinia-specific (also present in Y. pseudotuberculosis and Y. enterocolitica) Ysr141 (Yersinia small RNA 141). Ysr141 sRNA was shown to regulate the synthesis of the type III secretion system (T3SS) effector protein YopJ. The Yop-Ysc T3SS is a critical component of virulence for Yersinia species.
See also
Pathogenesis and immunity
In the urban and sylvatic (forest) cycles of Y. pestis, most of the spreading occurs between rodents and fleas. In the sylvatic cycle, the rodent is wild, but in the urban cycle, the rodent is primarily the brown rat. In addition, Y. pestis can spread from the urban environment and back. Transmission to humans is usually through the bite of infected fleas. If the disease has progressed to the pneumonic form, humans can spread the bacterium to others by coughing, vomiting, and possibly sneezing.
In reservoir hosts
Several species of rodents serve as the main reservoir for Y. pestis in the environment. In the steppes, the natural reservoir is believed to be principally the marmot. In the western United States, several species of rodents are thought to maintain Y. pestis. However, the expected disease dynamics have not been found in any rodent. A variety of species of rodents are known to have a variable resistance, which could lead to an asymptomatic carrier status. Evidence indicates fleas from other mammals have a role in human plague outbreaks.
The lack of knowledge of the dynamics of plague in mammal species is also true among susceptible rodents such as the black-tailed prairie dog (Cynomys ludovicianus), in which plague can cause colony collapse, resulting in a massive effect on prairie food webs. However, the transmission dynamics within prairie dogs does not follow the dynamics of blocked fleas; carcasses, unblocked fleas, or another vector could possibly be important, instead.
In other regions of the world, the reservoir of the infection is not clearly identified, which complicates prevention and early warning programs. One such example was seen in a 2003 outbreak in Algeria. The domestic house cat is susceptible to plague. Their symptoms are similar to those experienced by humans. Cats infected with plague can infect people through bites, scratches, coughs, or sneezes.
Vector
The transmission of Y. pestis by fleas is well characterized. Initial acquisition of Y. pestis by the vector occurs during feeding on an infected animal. Several proteins then contribute to the maintenance of the bacteria in the flea digestive tract, among them the hemin storage system and Yersinia murine toxin (Ymt). Although Ymt is highly toxic to rodents and was once thought to be produced to ensure reinfection of new hosts, it is important for the survival of Y. pestis in fleas.
The hemin storage system plays an important role in the transmission of Y. pestis back to a mammalian host. While in the insect vector, proteins encoded by hemin storage system genetic loci induce biofilm formation in the proventriculus, a valve connecting the midgut to the esophagus. Aggregation in the biofilm inhibits feeding, as a mass of clotted blood and bacteria forms (referred to as "Bacot's block"). Transmission of Y. pestis occurs during the futile attempts of the flea to feed. Ingested blood is pumped into the esophagus, where it dislodges bacteria lodged in the proventriculus and is regurgitated back into the host circulatory system.
In humans and other susceptible hosts
Pathogenesis due to Y. pestis infection of mammalian hosts is due to several factors, including an ability of these bacteria to suppress and avoid normal immune system responses such as phagocytosis and antibody production. Flea bites allow for the bacteria to pass the skin barrier. Y. pestis expresses the yadBC gene, which is similar to adhesins in other Yersinia species, allowing for adherence and invasion of epithelial cells. Y. pestis expresses a plasmin activator that is an important virulence factor for pneumonic plague and that might degrade on blood clots to facilitate systematic invasion. Many of the bacteria's virulence factors are anti-phagocytic in nature. Two important anti-phagocytic antigens, named F1 (Fraction 1) and V or LcrV, are both important for virulence. These antigens are produced by the bacterium at normal human body temperature. Furthermore, Y. pestis survives and produces F1 and V antigens while it is residing within white blood cells such as monocytes, but not in neutrophils. Natural or induced immunity is achieved by the production of specific opsonic antibodies against F1 and V antigens; antibodies against F1 and V induce phagocytosis by neutrophils.
In addition, the type-III secretion system (T3SS) allows Y. pestis to inject proteins into macrophages and other immune cells. These T3SS-injected proteins, called Yersinia outer proteins (Yops), include Yop B/D, which form pores in the host cell membrane and have been linked to cytolysis. The YopO, YopH, YopM, YopT, YopJ, and YopE are injected into the cytoplasm of host cells by T3SS into the pore created in part by YopB and YopD. The injected Yops limit phagocytosis and cell signaling pathways important in the innate immune system, as discussed below. In addition, some Y. pestis strains are capable of interfering with immune signaling (e.g., by preventing the release of some cytokines).
Y. pestis proliferates inside lymph nodes, where it is able to avoid destruction by cells of the immune system such as macrophages. The ability of Y. pestis to inhibit phagocytosis allows it to grow in lymph nodes and cause lymphadenopathy. YopH is a protein tyrosine phosphatase that contributes to the ability of Y. pestis to evade immune system cells. In macrophages, YopH has been shown to dephosphorylate p130Cas, Fyb (Fyn binding protein) SKAP-HOM and Pyk, a tyrosine kinase homologous to FAK. YopH also binds the p85 subunit of phosphoinositide 3-kinase, the Gab1, the Gab2 adapter proteins, and the Vav guanine nucleotide exchange factor.
YopE functions as a GTPase-activating protein for members of the Rho family of GTPases such as RAC1. YopT is a cysteine protease that inhibits RhoA by removing the isoprenyl group, which is important for localizing the protein to the cell membrane. It has been proposed that YopE and YopT may function to limit YopB/D-induced cytolysis. This might limit the function of YopB/D to create the pores used for Yop insertion into host cells and prevent YopB/D-induced rupture of host cells and release of cell contents that would attract and stimulate immune system responses.
YopJ is an acetyltransferase that binds to a conserved α-helix of MAPK kinases. YopJ acetylates MAPK kinases at serines and threonines that are normally phosphorylated during activation of the MAP kinase cascade. YopJ is activated in eukaryotic cells by interaction with target cell Phytic acid (IP6). This disruption of host cell protein kinase activity causes apoptosis of macrophages, and it has been proposed that this is important for the establishment of infection and for evasion of the host immune response. YopO is a protein kinase also known as Yersinia protein kinase A (YpkA). YopO is a potent inducer of human macrophage apoptosis.
Depending on which form of the plague the individual becomes infected with the plague develops different illness; however the plague overall effects the hosts cell’s ability to communicate with the immune system, hindering the body to bring phagocytic cells to the area of infection.
Immunity
A formalin-inactivated vaccine once was available in the United States for adults at high risk of contracting the plague until removal from the market by the Food and Drug Administration. It was of limited effectiveness and could cause severe inflammation. Experiments with genetic engineering of a vaccine based on F1 and V antigens are underway and show promise. However, bacteria lacking antigen F1 are still virulent, and the V antigens are sufficiently variable, such that vaccines composed of these antigens may not be fully protective. United States Army Medical Research Institute of Infectious Diseases (USAMRIID) have found that an experimental F1/V antigen-based vaccine protects crab-eating macaques but fails to protect African green monkey species. A systematic review by the Cochrane Collaboration found no studies of sufficient quality to make any statement on the efficacy of the vaccine.
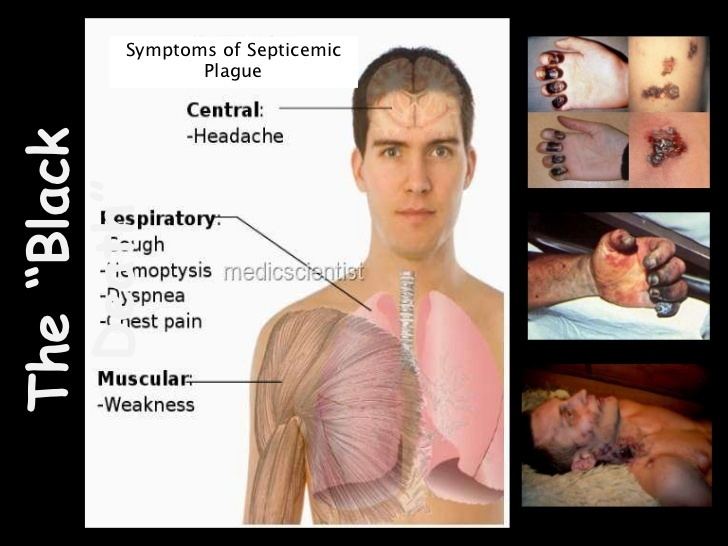
Symptoms and disease progression
Note: Patient may die before any symptoms appear
If this occurs with the classic buboes, this is considered primary, while secondary occurs after symptoms of bubonic or pneumonic infection. Since the bacteria are blood-borne, several organs can be affected, including the spleen and brain. The diffuse infection can cause an immunologic cascade to occur, leading to disseminated intravascular coagulation (DIC), which in turn results in bleeding and necrotic skin and tissue. Such a disseminated infection increases mortality to 22%.
With the exception of the buboes, the initial symptoms of plague are very similar to many other diseases, making diagnosis difficult.
ICD-9 codes for the diseases caused by Y. pestis:
Clinical determination
Gram's stains can confirm the presence of gram-negative rods, and in some cases the identification of the double-curved shape. An anti-F1 serology test can differentiate between different species of Yersinia, and Polymerase chain reaction (PCR) can be used to identify Y. pestis.
The protein H of the tail fiber of the bacteriophage Yersinia phage L-413C permits the differentiation between Y. pestis and Y. pseudotuberculosis", the gastro-intestinal corrolary (Kane et al.).
Treatment
The traditional second line treatment for Y. pestis has been streptomycin, chloramphenicol, tetracycline, and fluoroquinolones. There is also good evidence to support the use of doxycycline or gentamicin. Resistant strains have been isolated; treatment should be guided by antibiotic sensitivities where available. Antibiotic treatment alone is insufficient for some patients, who may also require circulatory, ventilator, or renal support.
In an emergency department setting, Harrison's Principles of Internal Medicine outlines the following treatment course. Antibiotics within the first 24 hours are very beneficial, with intravenous being preferred in pulmonary or advanced cases. Streptomycin or gentamicin are the first-line drugs, with chloramphenicol for critically ill patients, or rarely for suspected neuro-involvement.
Prevention
Plague vaccine is available, based on an attenuated type.
Plague of Justinian
During the mid-sixth century, the pandemic known as the Plague of Justinian wiped out roughly one third of the Byzantine Empire's population, creating major military and financial difficulties. Modern historians named this plague incident after the Eastern Roman Emperor Justinian I, who held power in the Byzantine capital of Constantinople at the time of the initial outbreak. The primary years of the plague were 541–542 AD, although plague returned throughout the Mediterranean basin in successive generations, until about 750.
The waves of disease had a major effect on the future course of European history. The plague's social and cultural impact during the Justinian period is comparable to that of the Black Death.
The most commonly accepted cause of the pandemic has been bubonic plague. A genetic study suggests the Plague of Justinian (and others from antiquity) arose from either now-extinct strains of Y. pestis, genetically distinct from the strain that broke out in the 14th-century pandemic, or from pathogens entirely unrelated to bubonic plague.
Role in Black Death
In the 1340s, Europe had a severe disease outbreak that began in the southern port cities in Italy and thereafter steadily spread northward, killing millions in its path. It often emptied entire villages of people and created mass hysteria. The death toll was so great, a very religious contingent thought God was punishing man for sin. When it finally reached England in 1348, it wound up destroying more than a third of the country's population. From that time on, the disease would recur repeatedly throughout Europe until the end of the 18th century, with the last outbreak in England occurring in 1666.
In 2000, Didier Raoult and others reported finding Y. pestis DNA by performing a "suicide PCR" on tooth pulp tissue from a 14th-century plague cemetery in Montpellier.
A study by an international team of researchers published in October 2010 confirmed Y. pestis was the cause of the Black Death and later epidemics on the entire European continent over a period of 400 years. The team used ancient DNA and proteins recovered from the bodies of plague victims buried in Hereford in England, in Saint-Laurent-de-la-Cabrerisse in France, and Bergen op Zoom in the Netherlands to identify the pathogen. They found two previously unknown, older strains of Y. pestis that had spread from China by two different routes, rather than the modern Y. p., orientalis and Y. p. medievalis.
Three biovars of Y. pestis were originally thought to correspond to one of the historical pandemics of bubonic plague. Biovar Y. p. antiqua is thought to correspond to the Plague of Justinian; it is not known whether this biovar also corresponds to earlier or smaller epidemics of bubonic plague, or whether these were even truly bubonic plague. Biovar Y. p. mediaevalis was formerly thought to correspond to the Black Death, while Biovar Y. p. orientalis was thought to correspond to the Third Pandemic and the majority of modern outbreaks of plague. However, calculations of Y. pestis' evolutionary age, found using the number of synonymous single-nucleotide polymorphisms (SNPs) in conjunction with molecular clock rates, date the emergence of the biovars prior to any of the historical epidemics due to the length of time needed to accumulate such mutations. Additional evidence against this hypothesis includes the fact that Y. p. mediaevalis is likely too young to have produced the Black Death due to its recent divergence from Y. p. orientalis.
Use in biological warfare
Y. pestis is potentially one of the first examples of biological warfare in (recorded) history, when in 1347, plague victims were catapulted by the Mongols over the city walls of Caffa, a town currently known as Feodosiya located in Crimea. Infected inhabitants may have fled to Italy, thus spreading the Black Death to Europe, though this is likely only one of a few routes that could have brought the plague from the east.
Y. pestis was used as a biological weapon in World War II, when on October 4, 1940, a Japanese airplane flying over Chushien, Chekiang Province, China, released rice and wheat plus rat fleas carrying Y. pestis. A second plane load was released three weeks later. These actions led to a local plague that killed 121 people.
Proposed use in biological warfare
Leon A. Fox from the U.S. Army Medical Corps had suggested a similar approach in 1933, proposing the possibility of infected rats being dropped from planes. U.S. Army Medical Corps Major Leon Fox published an article in 1933 in the magazine Military Surgeon writing: "Practically insurmountable difficulties prevent the use of biologic agents as effective weapons."
Recent events
In September 2009, the death of Malcolm Casadaban, a molecular genetics professor at the University of Chicago, was linked to his work on a weakened laboratory strain of Y. pestis. Hemochromatosis was hypothesised to be a predisposing factor in Casadaban's death from this attenuated strain used for research.
In 2012, researchers in Germany collected samples of Yersinia pestis from gravesites with a view to reconstructing the DNA of the bacterium. In 2015, Cell published results from a study of ancient graves. Plasmids of Y. pestis were detected in archaeological samples of the teeth of seven Bronze Age individuals, in the Afanasievo culture in Siberia, the Corded Ware culture in Estonia, the Sintashta culture in Russia, the Unetice culture in Poland and the Andronovo culture in Siberia.
June 8, 2015 Larimer County, CO Fatality confirmed By CDC as listed on the RSOE EDIS - Emergency and Disaster Information Service
September 8, 2016 Yersinia pestis bacterium identified from DNA in teeth from skeletons found at Crossrail Site, London showing the human remains were victims of the Great Plague http://www.crossrail.co.uk/news/articles/dna-of-bacteria-responsible-for-london-great-plague-of-1665-identified-for-first-time