
 | ||
Inflammation (from Latin inflammatio) is part of the complex biological response of body tissues to harmful stimuli, such as pathogens, damaged cells, or irritants, and is a protective response involving immune cells, blood vessels, and molecular mediators. The function of inflammation is to eliminate the initial cause of cell injury, clear out necrotic cells and tissues damaged from the original insult and the inflammatory process, and to initiate tissue repair.
Contents
- Cardinal signs
- Process of acute inflammation
- Vasodilation and increased permeability
- Plasma cascade systems
- Plasma derived mediators
- Cellular component
- Leukocyte extravasation
- Phagocytosis
- Cell derived mediators
- Morphologic patterns
- Inflammatory disorders
- Atherosclerosis
- Allergies
- Myopathies
- Leukocyte defects
- Pharmacological
- Cancer
- HIV and AIDS
- Resolution of inflammation
- Connection to depression
- Systemic effects
- Acute phase proteins
- Leukocyte numbers
- Systemic inflammation and obesity
- Systemic inflammation and overeating
- Outcomes
- Examples
- Diet and inflammation
- Derivation and computation
- Accuracy
- Disease prevention and causation
- Other factors that can influence inflammation
- Advantages and disadvantages of the DII
- Criticism and alternatives
- Exercise induced acute inflammation
- Post inflammatory muscle growth and repair
- Chronic inflammation and muscle loss
- Exercise as a treatment for inflammation
- Signal to noise theory
- References
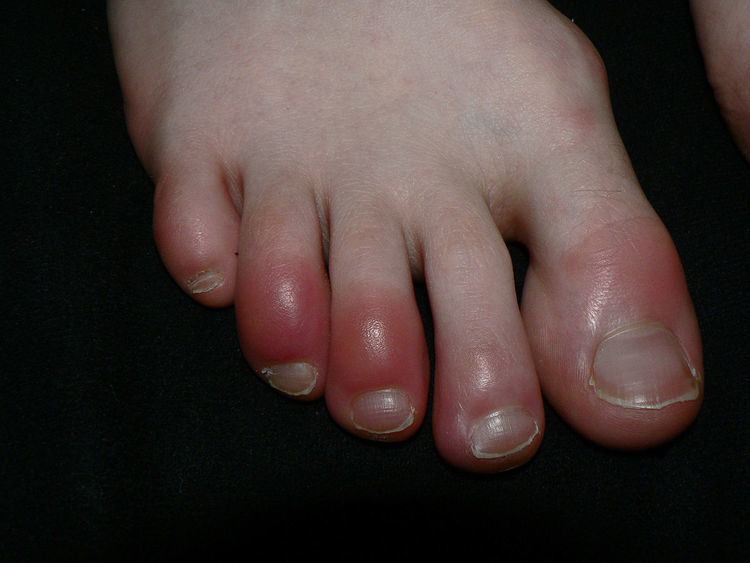
The classical signs of inflammation are heat, pain, redness, swelling, and loss of function. Inflammation is a generic response, and therefore it is considered as a mechanism of innate immunity, as compared to adaptive immunity, which is specific for each pathogen. Too little inflammation could lead to progressive tissue destruction by the harmful stimulus (e.g. bacteria) and compromise the survival of the organism. In contrast, chronic inflammation may lead to a host of diseases, such as hay fever, periodontitis, atherosclerosis, rheumatoid arthritis, and even cancer (e.g., gallbladder carcinoma). Inflammation is therefore normally closely regulated by the body.
Inflammation can be classified as either acute or chronic. Acute inflammation is the initial response of the body to harmful stimuli and is achieved by the increased movement of plasma and leukocytes (especially granulocytes) from the blood into the injured tissues. A series of biochemical events propagates and matures the inflammatory response, involving the local vascular system, the immune system, and various cells within the injured tissue. Prolonged inflammation, known as chronic inflammation, leads to a progressive shift in the type of cells present at the site of inflammation, such as mononuclear cells, and is characterized by simultaneous destruction and healing of the tissue from the inflammatory process.
Inflammation is not a synonym for infection. Infection describes the interaction between the action of microbial invasion and the reaction of the body's inflammatory response — the two components are considered together when discussing an infection, and the word is used to imply a microbial invasive cause for the observed inflammatory reaction. Inflammation on the other hand describes purely the body's immunovascular response, whatever the cause may be. But because of how often the two are correlated, words ending in the suffix -itis (which refers to inflammation) are sometimes informally described as referring to infection. For example, the word urethritis strictly means only "urethral inflammation", but clinical health care providers usually discuss urethritis as a urethral infection because urethral microbial invasion is the most common cause of urethritis.
It is useful to differentiate inflammation and infection as there are many pathological situations where inflammation is not driven by microbial invasion – for example, atherosclerosis, type III hypersensitivity, trauma, ischaemia. There are also pathological situations where microbial invasion does not result in classic inflammatory response—for example, parasitosis, eosinophilia.
Cardinal signs
Acute inflammation is a short-term process, usually appearing within a few minutes or hours and begins to cease upon the removal of the injurious stimulus. It involves a coordinated and systemic mobilisation response locally of various immune, endocrine and neurological mediators of acute inflammation. In a normal healthy response, it becomes activated, clears the pathogen and begins a repair process and then ceases. It is characterized by five cardinal signs:
An acronym that may be used to remember the key symptoms is "PRISH", for pain, redness, immobility (loss of function), swelling and heat.
The traditional names for signs of inflammation come from Latin:
The first four (classical signs) were described by Celsus (ca. 30 BC–38 AD), while loss of function was added later by Galen even though the attribution is disputed and the origination of the fifth sign has also been ascribed to Thomas Sydenham and Virchow.
Redness and heat are due to increased blood flow at body core temperature to the inflamed site; swelling is caused by accumulation of fluid; pain is due to the release of chemicals such as bradykinin and histamine that stimulate nerve endings. Loss of function has multiple causes.
Acute inflammation of the lung (usually caused in response to pneumonia) does not cause pain unless the inflammation involves the parietal pleura, which does have pain-sensitive nerve endings.
Process of acute inflammation
The process of acute inflammation is initiated by resident immune cells already present in the involved tissue, mainly resident macrophages, dendritic cells, histiocytes, Kupffer cells and mast cells. These cells possess surface receptors known as pattern recognition receptors (PRRs), which recognize (i.e., bind) two subclasses of molecules: pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs). PAMPs are compounds that are associated with various pathogens, but which are distinguishable from host molecules. DAMPs are compounds that are associated with host-related injury and cell damage.
At the onset of an infection, burn, or other injuries, these cells undergo activation (one of the PRRs recognize a PAMP or DAMP) and release inflammatory mediators responsible for the clinical signs of inflammation. Vasodilation and its resulting increased blood flow causes the redness (rubor) and increased heat (calor). Increased permeability of the blood vessels results in an exudation (leakage) of plasma proteins and fluid into the tissue (edema), which manifests itself as swelling (tumor). Some of the released mediators such as bradykinin increase the sensitivity to pain (hyperalgesia, dolor). The mediator molecules also alter the blood vessels to permit the migration of leukocytes, mainly neutrophils and macrophages, outside of the blood vessels (extravasation) into the tissue. The neutrophils migrate along a chemotactic gradient created by the local cells to reach the site of injury. The loss of function (functio laesa) is probably the result of a neurological reflex in response to pain.
In addition to cell-derived mediators, several acellular biochemical cascade systems consisting of preformed plasma proteins act in parallel to initiate and propagate the inflammatory response. These include the complement system activated by bacteria and the coagulation and fibrinolysis systems activated by necrosis, e.g. a burn or a trauma.
The acute inflammatory response requires constant stimulation to be sustained. Inflammatory mediators are short-lived and are quickly degraded in the tissue. Hence, acute inflammation begins to cease once the stimulus has been removed.
Vasodilation and increased permeability
As defined, acute inflammation is an immunovascular response to an inflammatory stimulus. This means acute inflammation can be broadly divided into a vascular phase that occurs first, followed by a cellular phase involving immune cells (more specifically myeloid granulocytes in the acute setting). The vascular component of acute inflammation involves the movement of plasma fluid, containing important proteins such as fibrin and immunoglobulins (antibodies), into inflamed tissue.
Upon contact with PAMPs, tissue macrophages and mastocytes release vasoactive amines such as histamine and serotonin, as well as eicosanoids such as prostaglandin E2 and leukotriene B4 to remodel the local vasculature. Macrophages and endothelial cells release nitric oxide. These mediators vasodilate and permeabilize the blood vessels, which results in the net distribution of blood plasma from the vessel into the tissue space. The increased collection of fluid into the tissue causes it to swell (edema). This exuded tissue fluid contain various antimicrobial mediators from the plasma such as complement, lysozyme, antibodies, which can immediately deal damage to microbes, and opsonise the microbes in preparation for the cellular phase. If the inflammatory stimulus is a lacerating wound, exuded platelets, coagulants, plasmin and kinins can clot the wounded area and provide haemostasis in the first instance. These clotting mediators also provide a structural staging framework at the inflammatory tissue site in the form of a fibrin lattice – as would construction scaffolding at a construction site – for the purpose of aiding phagocytic debridement and wound repair later on. Some of the exuded tissue fluid is also funnelled by lymphatics to the regional lymph nodes, flushing bacteria along to start the recognition and attack phase of the adaptive immune system.
Acute inflammation is characterized by marked vascular changes, including vasodilation, increased permeability and increased blood flow, which are induced by the actions of various inflammatory mediators. Vasodilation occurs first at the arteriole level, progressing to the capillary level, and brings about a net increase in the amount of blood present, causing the redness and heat of inflammation. Increased permeability of the vessels results in the movement of plasma into the tissues, with resultant stasis due to the increase in the concentration of the cells within blood – a condition characterized by enlarged vessels packed with cells. Stasis allows leukocytes to marginate (move) along the endothelium, a process critical to their recruitment into the tissues. Normal flowing blood prevents this, as the shearing force along the periphery of the vessels moves cells in the blood into the middle of the vessel.
Plasma cascade systems
Plasma-derived mediators
* non-exhaustive list
Cellular component
The cellular component involves leukocytes, which normally reside in blood and must move into the inflamed tissue via extravasation to aid in inflammation. Some act as phagocytes, ingesting bacteria, viruses, and cellular debris. Others release enzymatic granules that damage pathogenic invaders. Leukocytes also release inflammatory mediators that develop and maintain the inflammatory response. In general, acute inflammation is mediated by granulocytes, whereas chronic inflammation is mediated by mononuclear cells such as monocytes and lymphocytes.
Leukocyte extravasation
Various leukocytes, particularly neutrophils, are critically involved in the initiation and maintenance of inflammation. These cells must be able to move to the site of injury from their usual location in the blood, therefore mechanisms exist to recruit and direct leukocytes to the appropriate place. The process of leukocyte movement from the blood to the tissues through the blood vessels is known as extravasation, and can be broadly divided up into a number of steps:
- Leukocyte margination and endothelial adhesion: The white blood cells within the vessels which are generally centrally located move peripherally towards the walls of the vessels. Activated macrophages in the tissue release cytokines such as IL-1 and TNFα, which bind to their respective G protein-coupled receptors on the endothelial wall. Signal transduction induces the immediate expression of P-selectin on endothelial cell surfaces. This receptor binds weakly to carbohydrate ligands on the surface of leukocytes and causes them to "roll" along the endothelial surface as bonds are made and broken. Cytokines from injured cells induce the expression of E-selectin on endothelial cells, which functions similarly to P-selectin. Cytokines also induce the expression of integrin ligands such as ICAM-1 and VCAM-1 on endothelial cells, which mediate the adhesion and further slow leukocytes down. These weakly bound leukocytes are free to detach if not activated by chemokines produced in injured tissue. Activation increases the affinity of bound integrin receptors for ICAM-1 and VCAM-1 on the endothelial cell surface, firmly binding the leukocytes to the endothelium.
- Migration across the endothelium, known as transmigration, via the process of diapedesis: Chemokine gradients stimulate the adhered leukocytes to move between adjacent endothelial cells. The endothelial cells retract and the leukocytes pass through the basement membrane into the surrounding tissue using adhesion molecules such as ICAM-1.
- Movement of leukocytes within the tissue via chemotaxis: Leukocytes reaching the tissue interstitium bind to extracellular matrix proteins via expressed integrins and CD44 to prevent them from leaving the site. A variety of molecules behave as chemoattractants, for example, C3a or C5, and cause the leukocytes to move along a chemotactic gradient towards the source of inflammation.
Phagocytosis
Extravasated neutrophils in the cellular phase come into contact with microbes at the inflamed tissue. Phagocytes express cell-surface endocytic pattern recognition receptors (PRRs) that have affinity and efficacy against non-specific microbe-associated molecular patterns (PAMPs). Most PAMPs that bind to endocytic PRRs and initiate phagocytosis are cell wall components, including complex carbohydrates such as mannans and β-glucans, lipopolysaccharides (LPS), peptidoglycans, and surface proteins. Endocytic PRRs on phagocytes reflect these molecular patterns, with C-type lectin receptors binding to mannans and β-glucans, and scavenger receptors binding to LPS.
Upon endocytic PRR binding, actin-myosin cytoskeletal rearrangement adjacent to the plasma membrane occurs in a way that endocytoses the plasma membrane containing the PRR-PAMP complex, and the microbe. Phosphatidylinositol and Vps34-Vps15-Beclin1 signalling pathways have been implicated to traffic the endocytosed phagosome to intracellular lysosomes, where fusion of the phagosome and the lysosome produces a phagolysosome. The reactive oxygen species, superoxides and hypochlorite bleach within the phagolysosomes then kill microbes inside the phagocyte.
Phagocytic efficacy can be enhanced by opsonization. Plasma derived complement C3b and antibodies that exude into the inflamed tissue during the vascular phase bind to and coat the microbial antigens. As well as endocytic PRRs, phagocytes also express opsonin receptors Fc receptor and complement receptor 1 (CR1), which bind to antibodies and C3b, respectively. The co-stimulation of endocytic PRR and opsonin receptor increases the efficacy of the phagocytic process, enhancing the lysosomal elimination of the infective agent.
Cell-derived mediators
* non-exhaustive list
Morphologic patterns
Specific patterns of acute and chronic inflammation are seen during particular situations that arise in the body, such as when inflammation occurs on an epithelial surface, or pyogenic bacteria are involved.
Inflammatory disorders
Inflammatory abnormalities are a large group of disorders that underlie a vast variety of human diseases. The immune system is often involved with inflammatory disorders, demonstrated in both allergic reactions and some myopathies, with many immune system disorders resulting in abnormal inflammation. Non-immune diseases with etiological origins in inflammatory processes include cancer, atherosclerosis, and ischaemic heart disease.
A large variety of proteins are involved in inflammation, and any one of them is open to a genetic mutation which impairs or otherwise dysregulates the normal function and expression of that protein.
Examples of disorders associated with inflammation include:
Atherosclerosis
Atherosclerosis, formerly considered a bland lipid storage disease, actually involves an ongoing inflammatory response. Recent advances in basic science have established a fundamental role for inflammation in mediating all stages of this disease from initiation through progression and, ultimately, the thrombotic complications of atherosclerosis. These new findings provide important links between risk factors and the mechanisms of atherogenesis. Clinical studies have shown that this emerging biology of inflammation in atherosclerosis applies directly to human patients. Elevation in markers of inflammation predicts outcomes of patients with acute coronary syndromes, independently of myocardial damage. In addition, low-grade chronic inflammation, as indicated by levels of the inflammatory marker C-reactive protein, prospectively defines risk of atherosclerotic complications, thus adding to prognostic information provided by traditional risk factors. Moreover, certain treatments that reduce coronary risk also limit inflammation. In the case of lipid lowering with statins, this anti-inflammatory effect does not appear to correlate with reduction in low-density lipoprotein levels. These new insights into inflammation in atherosclerosis not only increase our understanding of this disease but also have practical clinical applications in risk stratification and targeting of therapy for this scourge of growing worldwide importance.
Allergies
An allergic reaction, formally known as type 1 hypersensitivity, is the result of an inappropriate immune response triggering inflammation, vasodilation, and nerve irritation. A common example is hay fever, which is caused by a hypersensitive response by mast cells to allergens. Pre-sensitised mast cells respond by degranulating, releasing vasoactive chemicals such as histamine. These chemicals propagate an excessive inflammatory response characterised by blood vessel dilation, production of pro-inflammatory molecules, cytokine release, and recruitment of leukocytes. Severe inflammatory response may mature into a systemic response known as anaphylaxis.
Other hypersensitivity reactions (type 2 and type 3) are mediated by antibody reactions and induce inflammation by attracting leukocytes that damage surrounding tissue.
Myopathies
Inflammatory myopathies are caused by the immune system inappropriately attacking components of muscle, leading to signs of muscle inflammation. They may occur in conjunction with other immune disorders, such as systemic sclerosis, and include dermatomyositis, polymyositis, and inclusion body myositis.
Leukocyte defects
Due to the central role of leukocytes in the development and propagation of inflammation, defects in leukocyte functionality often result in a decreased capacity for inflammatory defense with subsequent vulnerability to infection. Dysfunctional leukocytes may be unable to correctly bind to blood vessels due to surface receptor mutations, digest bacteria (Chédiak–Higashi syndrome), or produce microbicides (chronic granulomatous disease). In addition, diseases affecting the bone marrow may result in abnormal or few leukocytes.
Pharmacological
Certain drugs or exogenous chemical compounds are known to affect inflammation. Vitamin A deficiency causes an increase in inflammatory responses, and anti-inflammatory drugs work specifically by inhibiting the enzymes that produce inflammatory eicosanoids. Certain illicit drugs such as cocaine and ecstasy may exert some of their detrimental effects by activating transcription factors intimately involved with inflammation (e.g. NF-κB).
Cancer
Inflammation orchestrates the microenvironment around tumours, contributing to proliferation, survival and migration. Cancer cells use selectins, chemokines and their receptors for invasion, migration and metastasis. On the other hand, many cells of the immune system contribute to cancer immunology, suppressing cancer. Molecular intersection between receptors of steroid hormones, which have important effects on cellular development, and transcription factors that play key roles in inflammation, such as NF-κB, may mediate some of the most critical effects of inflammatory stimuli on cancer cells. This capacity of a mediator of inflammation to influence the effects of steroid hormones in cells, is very likely to affect carcinogenesis on the one hand; on the other hand, due to the modular nature of many steroid hormone receptors, this interaction may offer ways to interfere with cancer progression, through targeting of a specific protein domain in a specific cell type. Such an approach may limit side effects that are unrelated to the tumor of interest, and may help preserve vital homeostatic functions and developmental processes in the organism.
According to a review of 2009, recent data suggests that cancer-related inflammation (CRI) may lead to accumulation of random genetic alterations in cancer cells.
HIV and AIDS
It has long been recognized that infection with HIV is characterized not only by development of profound immunodeficiency but also by sustained inflammation and immune activation. A substantial body of evidence implicates chronic inflammation as a critical driver of immune dysfunction, premature appearance of aging-related diseases, and immune deficiency. Many now regard HIV infection not only as an evolving virus-induced immunodeficiency but also as chronic inflammatory disease. Even after the introduction of effective antiretroviral therapy (ART) and effective suppression of viremia in HIV-infected individuals, chronic inflammation persists. Animal studies also support the relationship between immune activation and progressive cellular immune deficiency: SIVsm infection of its natural nonhuman primate hosts, the sooty mangabey, causes high-level viral replication but limited evidence of disease. This lack of pathogenicity is accompanied by a lack of inflammation, immune activation and cellular proliferation. In sharp contrast, experimental SIVsm infection of rhesus macaque produces immune activation and AIDS-like disease with many parallels to human HIV infection.
Delineating how CD4 T cells are depleted and how chronic inflammation and immune activation are induced lies at the heart of understanding HIV pathogenesis––one of the top priorities for HIV research by the Office of AIDS Research, National Institutes of Health. Recent studies demonstrated that caspase-1-mediated pyroptosis, a highly inflammatory form of programmed cell death, drives CD4 T-cell depletion and inflammation by HIV. These are the two signature events that propel HIV disease progression to AIDS. Pyroptosis appears to create a pathogenic vicious cycle in which dying CD4 T cells and other immune cells (including macrophages and neutrophils) release inflammatory signals that recruit more cells into the infected lymphoid tissues to die. The feed-forward nature of this inflammatory response produces chronic inflammation and tissue injury. Identifying pyroptosis as the predominant mechanism that causes CD4 T-cell depletion and chronic inflammation, provides novel therapeutic opportunities, namely caspase-1 which controls the pyroptotic pathway. In this regard, pyroptosis of CD4 T cells and secretion of pro-inflmammatory cytokines such as IL-1β and IL-18 can be blocked in HIV-infected human lymphoid tissues by addition of the caspase-1 inhibitor VX-765, which has already proven to be safe and well tolerated in phase II human clinical trials. These findings could propel development of an entirely new class of “anti-AIDS” therapies that act by targeting the host rather than the virus. Such agents would almost certainly be used in combination with ART. By promoting “tolerance” of the virus instead of suppressing its replication, VX-765 or related drugs may mimic the evolutionary solutions occurring in multiple monkey hosts (e.g. the sooty mangabey) infected with species-specific lentiviruses that have led to a lack of disease, no decline in CD4 T-cell counts, and no chronic inflammation.
Resolution of inflammation
The inflammatory response must be actively terminated when no longer needed to prevent unnecessary "bystander" damage to tissues. Failure to do so results in chronic inflammation, and cellular destruction. Resolution of inflammation occurs by different mechanisms in different tissues. Mechanisms that serve to terminate inflammation include:
Connection to depression
There is evidence for a link between inflammation and depression. Inflammatory processes can be triggered by negative cognitions or their consequences, such as stress, violence, or deprivation. Thus, negative cognitions can cause inflammation that can, in turn, lead to depression.‹See TfD› In addition there is increasing evidence that inflammation can cause depression because of the increase of cytokines, setting the brain into a "sickness mode". Classical symptoms of being physically sick like lethargy show a large overlap in behaviors that characterize depression. Levels of cytokines tend to increase sharply during depressive episodes in manics and drop off during remission. Furthermore, it has been shown in clinical trials that anti-inflammatory medicines taken in addition to antidepressants not only significantly improves symptoms but also increases the proportion of subjects positively responding to treatment. Inflammations that lead to serious depression could be caused by common infections such as those caused by a virus, bacteria or even parasites.
Systemic effects
An infectious organism can escape the confines of the immediate tissue via the circulatory system or lymphatic system, where it may spread to other parts of the body. If an organism is not contained by the actions of acute inflammation it may gain access to the lymphatic system via nearby lymph vessels. An infection of the lymph vessels is known as lymphangitis, and infection of a lymph node is known as lymphadenitis. When lymph nodes cannot destroy all pathogens, the infection spreads further. A pathogen can gain access to the bloodstream through lymphatic drainage into the circulatory system.
When inflammation overwhelms the host, systemic inflammatory response syndrome is diagnosed. When it is due to infection, the term sepsis is applied, with the terms bacteremia being applied specifically for bacterial sepsis and viremia specifically to viral sepsis. Vasodilation and organ dysfunction are serious problems associated with widespread infection that may lead to septic shock and death.
Acute-phase proteins
Inflammation also induces high systemic levels of acute-phase proteins. In acute inflammation, these proteins prove beneficial; however, in chronic inflammation they can contribute to amyloidosis. These proteins include C-reactive protein, serum amyloid A, and serum amyloid P, which cause a range of systemic effects including:
Leukocyte numbers
Inflammation often affects the numbers of leukocytes present in the body:
Systemic inflammation and obesity
With the discovery of interleukins (IL), the concept of systemic inflammation developed. Although the processes involved are identical to tissue inflammation, systemic inflammation is not confined to a particular tissue but involves the endothelium and other organ systems.
Chronic inflammation is widely observed in obesity. The obese commonly have many elevated markers of inflammation, including:
Low-grade chronic inflammation is characterized by a two- to threefold increase in the systemic concentrations of cytokines such as TNF-α, IL-6, and CRP. Waist circumference correlates significantly with systemic inflammatory response. A predominant factor in this correlation is due to the autoimmune response triggered by adiposity, whereby immune cells may mistake fatty deposits for intruders. The body attacks fat similar to bacteria and fungi. When expanded fat cells leak or break open, macrophages mobilize to clean up and embed into the adipose tissue. Then macrophages release inflammatory chemicals, including TNF-α and (IL-6). TNF's primary role is to regulate the immune cells and induce inflammation. White blood cells then assist by releasing more cytokines. This link between adiposity and inflammation has been shown to produce 10–35% of IL-6 in a resting individual, and this production increases with increasing adiposity.
Loss of white adipose tissue reduces levels of inflammation markers. The association of systemic inflammation with insulin resistance and atherosclerosis is the subject of intense research.
In the obese mouse models, inflammation and macrophage-specific genes are upregulated in white adipose tissue (WAT). There were also signs of dramatic increase in circulating insulin level, adipocyte lipolysis and formation of multinucleate giant cells. The fat-derived protein called angiopoietin-like protein 2 (Angptl2) elevates in fat tissues. Higher than normal Angptl2 level in fat tissues develop inflammation as well as insulin and leptin resistance. Stored fat secretes Leptin to signal satiety. Leptin resistance plays a role in the process where appetite overrules the message of satiety. Angptl2 then starts an inflammatory cascade causing blood vessels to remodel and attract macrophages. Angptl2 is an adipocyte-derived inflammatory mediator linking obesity to systemic insulin resistance. It is possible that, as an inflammatory marker, leptin responds specifically to adipose-derived inflammatory cytokines.
C-reactive protein (CRP) is generated at a higher level in obese people. It raises when there is inflammation throughout the body. Mild elevation in CRP increase risk of heart attacks, strokes, high blood pressure, muscle weakness and fragility.
Systemic inflammation and overeating
Hyperglycemia induces IL-6 production from endothelial cells and macrophages. Meals high in saturated fat, as well as meals high in calories have been associated with increases in inflammatory markers. While the inflammatory responses are acute and arise in response to overeating, the response may become chronic if the overeating is chronic.
Bente Klarlund Pedersen wrote in 2013 that interstitial abdominal adiposity (also referred to as accumulated intra-abdominal fat) is a major and possibly primary factor in increasing systemic risk for multiple inflammatory diseases. However, she indicates that this is mediated via TNF-α, rather than Interleukin 6, which she identified as a myokine in 2003. Dr. Pedersen concludes: "Until the beginning of this millennium, it was commonly thought that the increase in IL-6 during exercise was a consequence of an immune response due to local damage in the working muscles and it was hypothesized that macrophages were responsible for this increase. However, an early study demonstrated that IL-6 mRNA in monocytes did not increase as a result of exercise."
Dr. Pedersen continues (in the same article): "Patients with diabetes have a high protein expression of TNF-α in skeletal muscle and increased TNF-α levels in plasma, and it is likely that adipose tissue, which produces TNF-α, is the main source of the circulating TNF-α. In vitro studies demonstrate that TNF-α has direct inhibitory effects on insulin signaling. In addition, TNF-α infusion in healthy humans induces insulin resistance in skeletal muscle, without an effect on EGP [endogenous glucose production]. It has also been proposed that TNF-α causes insulin resistance indirectly in vivo by increasing the release of FFAs from adipose tissue. TNF-α increases lipolysis in human and 3T3-L1 adipocytes. However, TNF-α has no effect on muscle fatty acid oxidation, but increases fatty acid incorporation into diacylglycerol, which may be involved in the development of the TNF-α-induced insulin resistance in skeletal muscle. In addition, evidence suggests that TNF-α plays a direct role in linking insulin resistance to vascular disease. Moreover, in CVDs, activated immune cells also play major roles, particularly in the etiology of atherosclerosis. Importantly, also tumor initiation, promotion, and progression is stimulated by systemic elevation of proinflammatory cytokines."
Outcomes
The outcome in a particular circumstance will be determined by the tissue in which the injury has occurred and the injurious agent that is causing it. Here are the possible outcomes to inflammation:
- Resolution
The complete restoration of the inflamed tissue back to a normal status. Inflammatory measures such as vasodilation, chemical production, and leukocyte infiltration cease, and damaged parenchymal cells regenerate. In situations where limited or short-lived inflammation has occurred this is usually the outcome. - Fibrosis
Large amounts of tissue destruction, or damage in tissues unable to regenerate, cannot be regenerated completely by the body. Fibrous scarring occurs in these areas of damage, forming a scar composed primarily of collagen. The scar will not contain any specialized structures, such as parenchymal cells, hence functional impairment may occur. - Abscess formation
A cavity is formed containing pus, an opaque liquid containing dead white blood cells and bacteria with general debris from destroyed cells. - Chronic inflammation
In acute inflammation, if the injurious agent persists then chronic inflammation will ensue. This process, marked by inflammation lasting many days, months or even years, may lead to the formation of a chronic wound. Chronic inflammation is characterised by the dominating presence of macrophages in the injured tissue. These cells are powerful defensive agents of the body, but the toxins they release (including reactive oxygen species) are injurious to the organism's own tissues as well as invading agents. As a consequence, chronic inflammation is almost always accompanied by tissue destruction.
Examples
Inflammation is usually indicated by adding the suffix "itis", as shown below. However, some conditions such as asthma and pneumonia do not follow this convention. More examples are available at list of types of inflammation.
Diet and inflammation
The Dietary Inflammatory Index (DII) is a score (number) that describes the potential of diet to modulate systemic inflammation within the body. As stated chronic inflammation is linked to most chronic diseases including arthritis, many types of cancer, cardiovascular diseases, inflammatory bowel diseases, and diabetes.
It has been known for many decades both that inflammation is linked to health and that diet is a major determinant of inflammation. Until the invention of the DII by scientists led by James R. Hébert at the Statewide South Carolina Cancer Prevention and Control Program in the University of South Carolina, no indicator of diet-related inflammation had ever been devised. The DII is unique among dietary indices in that it is based on results of studies published in the peer-reviewed literature and not any of the three methods typically used to create a dietary index; i.e., Basing it on 1) a particular cuisine or foodway (e.g., the Mediterranean, Macrobiotic or South Asian diet); 2) dietary guidelines (e.g., the Healthy Eating Index, which is based on the US Dietary Guidelines); or 3) results from a particular study (which reflect biases related to sample selection, response set biases such as social desirability). By contrast, the DII: 1) is grounded in peer-reviewed literature focusing specifically on inflammation (thus obviating idiosyncrasies of specific dietary assessment methods and limitations of exposure differences within populations); 2) can be adapted to virtually any dietary assessment method capable of providing estimates of nutrient intake; and 3) has been standardized to dietary intake from representative populations around the world, thus facilitating easy quantitative comparisons across studies.
Since its invention in 2004 and the publication of a now-defunct earlier-version of the DII in December 2009, the DII went through a major overhaul, resulting in a new version that replaced the old version, published first on-line in 2013 and then in hard copy in 2014. This new version is superior to the earlier one in a number of ways: 1) it is based on a larger, more robust literature (see Derivation/ Computation) of 1,943 (vs. 927) articles that were reviewed and scored; 2) creation of a global dataset based on intake measures from populations in 11 countries around the world, thus allowing comparison of results from studies conducted anywhere in the world; and 3) scoring such that lower, more negative, scores indicate anti-inflammation, while higher, more positive, scores are more pro-inflammatory.
This new, improved version of the DII has been subjected to construct validation, which tested (and subsequently confirmed) its ability to predict blood levels of inflammatory markers. The first of these tests showed the DII could predict repeat measures of c-reactive protein (CRP) from the Seasonal Variation of Blood Cholesterol, an intensive follow-up study of 559 adult men and women conducted in Massachusetts. The second, based on data from 2,567 women across the USA participating in the Women’s Health Initiative Observational Study from 1993–1998, showed the DII’s ability to predict levels of interleukin (IL)-6 and tumor necrosis factor-alpha receptor-2 (TNFα-R2) and a combined inflammatory biomarker score (consisting of IL-6, TNFα-R2, and CRP). Subsequently, the DII has been shown to have positive associations with IL-6 and homocysteine in the Asklepios Study in Belgium, CRP among police officers in Buffalo, NY, USA and inversely correlated with SNP rs2243250 in IL-4 among controls in the Bellvitge Colorectal Cancer Study in Spain.
Derivation and computation
Briefly, based on a search of the literature from 1950 to the end of 2010, 45 food parameters were identified among foods, nutrients, and other food components that were associated with 6 plasma inflammatory markers (IL-1β, IL-4, IL-6, IL-10, TNF-α and CRP). A specific article inflammatory effect score for each food parameter on the basis of the literature review and taking into account the quality and number of published papers (1,943 articles were reviewed and scored). For each study participant in a study, the dietary data is first linked to a global database that was developed based on 11 datasets from around the world (US – NHANES; UK – The National Diet & Nutrition Survey; Bahrain – National Nutrition Survey for Adult Bahrainis; Mexico – Mexican National Health and Nutrition Survey; Australia – National Nutrition Survey; South Korea – KNHANES; Taiwan – Nutrition and Health Survey in Taiwan; India – Indian Diet Study; New Zealand – National Nutrition Survey) and thus provides a robust estimate of the mean and the standard deviation of these 45 parameters.2 Each subject’s exposure relative to the “standard global mean” is expressed as a z-score that is derived by subtracting the “standard global mean” from the amount reported, and dividing this value by its standard deviation. To minimize the effect of “right skewing”, this value was then converted to a centered percentile score. The subject’s DII score was computed by multiplying these values by the specific article inflammatory effect score for each food parameter and then summing together all these 45 values according to the following formula, DII=b1*n1+b2*n2...........b45*n45, where bi refers to the literature-derived inflammatory effect score for each of the evaluated food parameter and ni refers to the food parameter-specific centered percentile, which were derived from the dietary data, per each i from 1 to 45.
Accuracy
The DII is uniquely and strongly based on empirical evidence available in the peer-reviewed literature on the relationships between dietary factors and important inflammatory markers (see Derivation/ Computation). It is important to note, however, that, as an index that superimposes on a method of dietary assessment, the DII is dependent on the accuracy of the dietary assessment method used. The DII cannot make up for deficiencies/ inaccuracies in measuring dietary intake.
Disease prevention and causation
Besides studies focusing on markers of inflammation in blood, the DII has been associated with a large number of actual health outcomes including:
Other factors that can influence inflammation
Besides dietary factors, physical activity, fitness, sleep, shift work, and stress can affect inflammation. Also, there are auto-immune and other conditions that can influence inflammation. It has been shown that these other factors also may be related to the DII.
Advantages and disadvantages of the DII
Advantages of the DII over other indices include that it: 1) focuses on a particular and very important mechanism of how dietary components affect health through inflammation; 2) is grounded in peer-reviewed literature focusing specifically on inflammatory markers known to affect health well-being; 3) can be adapted to virtually any dietary assessment method that provides estimates of nutrient intake; and 4) is standardized to dietary intake from representative populations around the world, thus facilitating easy quantitative comparisons across studies.
Disadvantages include that: 1) it is dependent on the quality of the dietary assessment upon which the computation is based (it cannot make up for deficiencies in the methods of obtaining the dietary data); and 2) it is not focused specifically on other effects of diet, though it is correlated (r (correlation coefficient) around │0.50│) with other diet indices that purport to assess diet quality.
Criticism and alternatives
Criticisms include that the DII is not up to date. It must be noted that though the index was refined by the inclusion of three more years (from 2008 through 2010) of publications and more than doubling of the literature base, no major change in the overall thrust of the literature base was evident.
Exercise-induced acute inflammation
Acute inflammation of the muscle cells, as understood in exercise physiology, can result after induced eccentric and concentric muscle training. Participation in eccentric training and conditioning, including resistance training and activities that emphasize eccentric lengthening of the muscle including downhill running on a moderate to high incline can result in considerable soreness within 24 to 48 hours, even though blood lactate levels, previously thought to cause muscle soreness, were much higher with level running. This delayed onset muscle soreness (DOMS) results from structural damage to the contractile filaments and z-disks, which has been noted especially in marathon runners whose muscle fibers revealed remarkable damage to the muscle fibers after both training and marathon competition. The onset and timing of this gradient damage to the muscle parallels the degree of muscle soreness experienced by the runners.
Z-disks are the point of contact for the contractile proteins. They provide structural support for transmission of force when muscle fibers are activated to shorten. However, in marathon runners and those who subscribe to the overload principle to enhance their muscles, show moderate Z-disk streaming and major disruption of thick and thin filaments in parallel groups of sarcomeres as a result of the force of eccentric actions or stretching of tightened muscle fibers.
This disruption of muscle fibers triggers white blood cells to increase following induced muscle soreness, leading to the inflammatory response observation from induced muscle soreness. Elevations in plasma enzymes, myoglobinemia, and abnormal muscle histology and ultrastructure are concluded to be associated with inflammatory response. High tension in the contractile-elastic system of muscle results in structural damage to the muscle fiber and plasmalemma and its epimysium, perimysium, and/or endomysium. The mysium damage disrupts calcium homeostasis in injured fibers and fiber bundles, resulting in necrosis that peaks about 48 hours after exercise. The products of macrophage activity and intracellular contents (such as histamines, kinins, and K+) accumulate outside cells. These substances then stimulate free nerve endings in the muscle; a process that appears accentuated by eccentric exercise, in which large forces are distributed over a relatively small cross-sectional area of the muscle.
Post-inflammatory muscle growth and repair
There is a known relationship between inflammation and muscle growth. For instance, high doses of anti-inflammatory medicines (e.g., NSAIDs) are able to blunt muscle growth. Cold therapy has been shown to negatively affect muscle growth as well. Reducing inflammation results in decreased macrophage activity and lower levels of IGF-1 Acute effects of cold therapy on training adaptations show reduced satellite cell proliferation. Long term effects include less muscular hypertrophy and an altered cell structure of muscle fibers.
It has been further theorized that the acute localized inflammatory responses to muscular contraction during exercise, as described above, are a necessary precursor to muscle growth. As a response to muscular contractions, the acute inflammatory response initiates the breakdown and removal of damaged muscle tissue. Muscles can synthesize cytokines in response to contractions, such that the cytokines interleukin-1 beta (IL-1β), TNF-α, and IL-6 are expressed in skeletal muscle up to 5 days after exercise.
In particular, the increase in levels of IL-6 (interleukin 6), a myokine, can reach up to one hundred times that of resting levels. Depending on volume, intensity, and other training factors, the IL-6 increase associated with training initiates about 4 hours after resistance training and remains elevated for up to 24 hours.
These acute increases in cytokines, as a response to muscle contractions, help initiate the process of muscle repair and growth by activating satellite cells within the inflamed muscle. Satellite cells are crucial for skeletal muscle adaptation to exercise. They contribute to hypertrophy by providing new myonuclei and repair damaged segments of mature myofibers for successful regeneration following injury- or exercise-induced muscle damage; high-level powerlifters can have up to 100% more satellite cells than untrained controls.
A rapid and transient localization of the IL-6 receptor and increased IL-6 expression occurs in satellite cells following contractions. IL-6 has been shown to mediate hypertrophic muscle growth both in vitro and in vivo. Unaccustomed exercise can increase IL-6 by up to sixfold at 5 hours post-exercise and threefold 8 days after exercise. Also telling is the fact that NSAIDs can decrease satellite cell response to exercise, thereby reducing exercise-induced protein synthesis.
The increase in cytokines (myokines) after resistance exercise coincides with the decrease in levels of myostatin, a protein that inhibits muscle differentiation and growth. The cytokine response to resistance exercise and moderate-intensity running occur differently, with the latter causing a more prolonged response, especially at the 12–24 hour mark.
Developing research has demonstrated that many of the benefits of exercise are mediated through the role of skeletal muscle as an endocrine organ. That is, contracting muscles release multiple substances known as myokines, including but not limited to those cited in the above description, which promote the growth of new tissue, tissue repair, and various anti-inflammatory functions, which in turn reduce the risk of developing various inflammatory diseases. The new view that muscle is an endocrine organ is transforming our understanding of exercise physiology and with it, of the role of inflammation in adaptation to stress.
Chronic inflammation and muscle loss
Both chronic and extreme inflammation are associated with disruptions of anabolic signals initiating muscle growth. Chronic inflammation has been implicated as part of the cause of the muscle loss that occurs with aging. Increased protein levels of myostatin have been described in patients with diseases characterized by chronic low-grade inflammation. Increased levels of TNF-α can suppress the AKT/mTOR pathway, a crucial pathway for regulating skeletal muscle hypertrophy, thereby increasing muscle catabolism. Cytokines may antagonize the anabolic effects of insulin-like growth factor 1 (IGF-1). In the case of sepsis, an extreme whole body inflammatory state, the synthesis of both myofibrillar and sarcoplasmic proteins are inhibited, with the inhibition taking place preferentially in fast-twitch muscle fibers. Sepsis is also able to prevent leucine from stimulating muscle protein synthesis. In animal models, when inflammation is created, mTOR loses its ability to be stimulated by muscle growth.
Exercise as a treatment for inflammation
Regular physical activity is reported to decrease markers of inflammation, although the correlation is imperfect and seems to reveal differing results contingent upon training intensity. For instance, while baseline measurements of circulating inflammatory markers do not seem to differ greatly between healthy trained and untrained adults, long-term training may help reduce chronic low-grade inflammation. On the other hand, levels of the anti-inflammatory myokine IL-6 (interleukin 6) remained elevated longer into the recovery period following an acute bout of exercise in patients with inflammatory diseases, relative to the recovery of healthy controls. It may well be that low-intensity training can reduce resting pro-inflammatory markers (CRP, IL-6), while moderate-intensity training has milder and less-established anti-inflammatory benefits. There is a strong relationship between exhaustive exercise and chronic low-grade inflammation. Marathon running may enhance IL-6 levels as much as 100 times over normal and increases total leuckocyte count and neturophil mobilization.
Regarding the above, IL-6 had previously been classified as a proinflammatory cytokine. Therefore, it was first thought that the exercise-induced IL-6 response was related to muscle damage. However, it has become evident that eccentric exercise is not associated with a larger increase in plasma IL-6 than exercise involving concentric “nondamaging” muscle contractions. This finding clearly demonstrates that muscle damage is not required to provoke an increase in plasma IL-6 during exercise. As a matter of fact, eccentric exercise may result in a delayed peak and a much slower decrease of plasma IL-6 during recovery.
Recent work has shown that both upstream and downstream signalling pathways for IL-6 differ markedly between myocytes and macrophages. It appears that unlike IL-6 signalling in macrophages, which is dependent upon activation of the NFκB signalling pathway, intramuscular IL-6 expression is regulated by a network of signalling cascades, including the Ca2+/NFAT and glycogen/p38 MAPK pathways. Thus, when IL-6 is signalling in monocytes or macrophages, it creates a pro-inflammatory response, whereas IL-6 activation and signalling in muscle is totally independent of a preceding TNF-response or NFκB activation, and is anti-inflammatory.
Several studies show that markers of inflammation are reduced following longer-term behavioural changes involving both reduced energy intake and a regular program of increased physical activity, and that, in particular, IL-6 was miscast as an inflammatory marker. For example, the anti-inflammatory effects of IL-6 have been demonstrated by IL-6 stimulating the production of the classical anti-inflammatory cytokines IL-1ra and IL-10. As such, individuals pursuing exercise as a means to treat the causal factors underlying chronic inflammation are pursuing a course of action strongly supported by current research, as an inactive lifestyle is strongly associated with the development and progression of multiple inflammatory diseases. Note that cautions regarding over-exertion may apply in certain cases, as discussed above, though this concern rarely applies to the general population.
Signal-to-noise theory
Given that localized acute inflammation is a necessary component for muscle growth, and that chronic low-grade inflammation is associated with a disruption of anabolic signals initiating muscle growth, it has been theorized that a signal-to-noise model may best describe the relationship between inflammation and muscle growth. By keeping the "noise" of chronic inflammation to a minimum, the localized acute inflammatory response signals a stronger anabolic response than could be achieved with higher levels of chronic inflammation.