
 | ||
Eicosanoids are signaling molecules made by the enzymatic or non-enzymatic oxidation of arachidonic acid or other polyunsaturated fatty acids (PUFAs) that are, like arachidonic acid, 20 carbon units in length. They are a sub-category of oxylipins, i.e. oxidized fatty acids of diverse carbon units in length but are further distinguished from other oxylipins by their overwhelming importance as cell signaling molecules. Eicosanoids function in diverse physiological and pathological systems including those: mounting or inhibiting inflammation, allergy, fever and other immune responses; influencing the abortion of pregnancy and normal childbirth; contributing to the perception of pain; regulating cell growth; controlling blood pressure; and modulating the regional flow of blood to tissues. In performing these tasks, eicosanoids most often act as autocrine signaling agents to impact their cells of origin or as paracrine signaling agents to impact cells near to their cells of origin. However, they can also act as endocrine agents to control the function of distant cells.
Contents
- Fatty acid sources
- Abbreviation
- Classic eicosanoids
- Nonclassic eicosanoids
- Biosyntheses
- Fatty acid mobilization
- Peroxidation and reactive oxygen species
- Prostanoid pathways
- Hydroxyeicosatetraenoate HETE and leukotriene LT pathways
- Epoxyeicosanoid pathway
- Function pharmacology and clinical significance
- Prostanoids
- Cyclopentenone prostaglandins
- HETEs and oxo ETEs
- leukotrienes
- Lipoxins and epi lipoxins
- Eoxins
- Resolvin metabolites of eicosapentaenoic acid
- Other metabolites of eicosapentaenoic acid
- Epoxyeicosanoids
- The 3 and 6 series
- Mechanisms of 3 action
- Role in inflammation
- History
- References
There are multiple subfamilies of eicosanoids, including most prominently the prostaglandins, thromboxanes, leukotrienes, lipoxins, resolvins, and eoxins but also others as noted in the following Nomenclature section. For each subfamily, there is the potential to have at least 5 separate series of metabolites, two series derived from ω-6 PUFAs (arachidonic and dihomo-gamma-linolenic acids), one series derived from the ω-3 PUFA (eicosapentaenoic acid), and one series derived from the ω-9 PUFA (mead acid). This subfamily distinction is important. Mammals, including humans, are unable to convert ω-6 into ω-3 PUFA. In consequence, tissue levels of the ω-6 and ω-3 PUFAs and their corresponding eicosanoid metabolites link directly to the amount of dietary ω-6 versus ω-3 PUFAs consumed. Since certain of the ω-6 and ω-3 PUFA series of metabolites have almost diametrically opposing physiological and pathological activities, it has often been suggested that the deleterious consequences associated with the consumption of ω-6 PUFA-rich diets reflects excessive production and activities of ω-6 PUFA-derived eicosanoids while the beneficial effects associated with the consumption of ω-3 PUFA-rich diets reflect the excessive production and activities of ω-3 PUFA-derived eicosanoids. In this view, the opposing effects of ω-6 PUFA-derived and ω-3 PUFA-derived eicosanoids on key target cells underlie the detrimental and beneficial effects of ω-6 and ω-3 PUFA-rich diets on inflammation and allergy reactions, atherosclerosis, hypertension, cancer growth, and a host of other processes.
Fatty acid sources
"Eicosanoid" (eicosa-, Greek for "twenty"; see icosahedron) is the collective term for straight-chain polyunsaturated fatty acids (PUFAs) of 20 carbon units in length that have been metabolized or otherwise converted to oxygen-containing products. The PUFA precursors to the eicosanoids include:
Abbreviation
A particular eicosanoid is denoted by a four-character abbreviation, composed of:
The stereochemistry of the eicosanoid products formed may differ among the pathways. For prostaglandins, this is often indicated by Greek letters (e.g. PGF2α versus PGF2β). For hydroperoxy and hydroxy eicosanoids an S or R designates the chirality of their substituents (e.g. 5S-hydroxy-eicosateteraenoic acid [also termed 5(S)-, 5S-hydroxy-, and 5(S)-hydroxy-eicosatetraenoic acid] is given the trivial names of 5S-HETE, 5(S)-HETE, 5S-HETE, or 5(S)-HETE). Since eicosanoid-forming enzymes commonly make S isomer products either with marked preference or essentially exclusively, the use of S/R designations has often been dropped (e.g. 5S-HETE is 5-HETE). Nonetheless, certain eicosanoid-forming pathways do form R isomers and their S versus R isomeric products can exhibit dramatically different biological activities. Failing to specify S/R isomers can be misleading. Here, all hydroperoxy and hydroxy substituents have the S configuration unless noted otherwise.
Classic eicosanoids
Current usage limits the term eicosanoid to:
Hydroxyeicosatetraenoic acids, leukotrienes, eoxins and prostanoids are sometimes termed "classic eicosanoids"
Nonclassic eicosanoids
In contrast to the classic eicosanoids, several other classes of PUFA metabolites have been termed 'novel', 'eicosanoid-like' or 'nonclassic eicosanoids'. These included the following classes:
Metabolism of eicosapentaenoic acid to HEPEs, leukotrienes, prostanoids, and epoxyeicosatetraenoic acids as well as the metabolism of dihomo-gamma-linolenic acid to prostanoids and mead acid to 5(S)-hydroxy-6E,8Z,11Z-eicosatrienoic acid (5-HETrE), 5-oxo-6,8,11-eicosatrienoic acid (5-oxo-ETrE), LTA3, and LTC3 involve the same enzymatic pathways that make their arachidonic acid-derived analogs.
Biosyntheses
Eicosanoids typically are not stored within cells but rather synthesized as required. They derive from the fatty acids that make up the cell membrane and nuclear membrane. These fatty acids must be released from their membrane sites and then metabolized initially to products which most often are further metabolized through various pathways to make the large array of products we recognize as bioactive eicosanoids.
Fatty acid mobilization
Eicosanoid biosynthesis begins when a cell is activated by mechanical trauma, ischemia, other physical perturbations, attack by pathogens, or stimuli made by nearby cells, tissues, or pathogens such as chemotactic factors, cytokines, growth factors, and even certain eicosanoids. The activated cells then mobilize enzymes, termed phospholipase A2's (PLA2s), capable of releasing ω-6 and ω-3 fatty acids from membrane storage. These fatty acids are bound in ester linkage to the SN2 position of membrane phospholipids; PLA2s act as esterases to release the fatty. There are several classes of PLA2s with type IV cytosolic PLA2s (cPLA2s) appearing to be responsible for releasing the fatty acids under many conditions of cell activation. The cPLA2s act specifically on phospholipids that contain AA, EPA or GPLA at their SN2 position. Interestingly, cPLA2 may also release the lysophospholipid that becomes platelet-activating factor.
Peroxidation and reactive oxygen species
Next, the free fatty acid is oxygenated along any of several pathways; see the Pathways table. The eicosanoid pathways (via lipoxygenase or COX) add molecular oxygen (O2). Although the fatty acid is symmetric, the resulting eicosanoids are chiral; the oxidations proceed with high stereoselectivity (enzymatic oxidations are considered practically stereospecific).
Four families of enzymes initiate or contribute to the initiation of the catalysis of fatty acids to eicosanoids:
Two different enzymes may act in series on a PUFA to form more complex metabolites. For example, ALOX5 acts with ALOX12 or aspirin-treated COX-2 to metabolize arachidonic acid to lipoxins and with cytochrome P450 monooxygenase(s), bacterial cytochrome P450 (in infected tissues), or aspirin-treated COX2 to metabolize eicosapentaenoic acid to the E series resolvins (RvEs) (see Specialized pro-resolving mediators). When this occurs with enzymes located in different cell types and involves the transfer of one enzyme's product to a cell which uses the second enzyme to make the final product it is referred to as transcellular metabolism or transcellular biosynthesis.
The oxidation of lipids is hazardous to cells, particularly when close to the nucleus. There are elaborate mechanisms to prevent unwanted oxidation. COX, the lipoxygenases, and the phospholipases are tightly controlled—there are at least eight proteins activated to coordinate generation of leukotrienes. Several of these exist in multiple isoforms.
Oxidation by either COX or lipoxygenase releases reactive oxygen species (ROS) and the initial products in eicosanoid generation are themselves highly reactive peroxides. LTA4 can form adducts with tissue DNA. Other reactions of lipoxygenases generate cellular damage; murine models implicate 15-lipoxygenase in the pathogenesis of atherosclerosis. The oxidation in eicosanoid generation is compartmentalized; this limits the peroxides' damage. The enzymes that are biosynthetic for eicosanoids (e.g., glutathione-S-transferases, epoxide hydrolases, and carrier proteins) belong to families whose functions are involved largely with cellular detoxification. This suggests that eicosanoid signaling might have evolved from the detoxification of ROS.
The cell must realize some benefit from generating lipid hydroperoxides close-by its nucleus. PGs and LTs may signal or regulate DNA-transcription there; LTB4 is ligand for PPARα. (See diagram at PPAR).
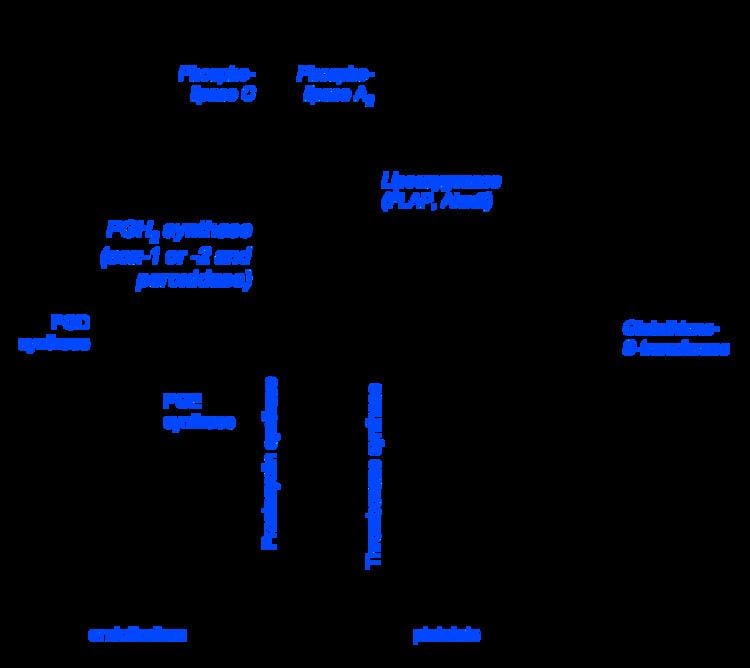
Prostanoid pathways
Both COX1 and COX2 (also termed prostaglandin-endoperoxide synthase-1 (PTGS1) and PTGS2, respectively) metabolize arachidonic acid by adding molecular O2 between carbons 9 and 11 to form an endoperoxide bridge between these two carbons, adding molecular O2 to carbon 15 to yield a 15-hydroperoxy product, creating a carbon-carbon bond between carbons 8 and 12 to create a cyclopentane ring in the middle of the fatty acid, and in the process making PGG2, a product that has two fewer double bonds than arachidonic acid. The 15-hydrpperoxy residue of PGG2 is then reduced to a 15-hydroxyl residue thereby forming PGH2. PGH2 is the parent prostanoid to all other postanoids. It is metabolized by (see diagram in Prostanoids: a) the Prostaglandin E synthase pathway in which any one of three isozymes, PTGES, PTGES2, or PTGES3, convert PGH2 to PGE2 (subsequent products of this pathway include PGA2 and PGB2 (see Prostanoid#Biosynthesis); b) PGF synthase which converts PGH2 to PGF2α; c) Prostaglandin D2 synthase which converts PGH2 to PGD2 (subsequent products in this pathway include 15-dPGJ2 (see Cyclopentenone prostaglandin); d) thromboxane synthase which converts PGH2 to TXA2 (subsequent products in this pathway include TXB2); and e) Prostacyclin synthase which converts PGH2 to PGI2 (subsequent products in this pathway include 6-keto-PGFα. These pathways have been shown or in some cases presumed to metabolize eicosapentaenoic acid to eicosanoid analogs of the sited products that have three rather than two double bonds and therefore contain the number 3 in place of 2 attached to their names (e.g. PGE3 instead of PGE2).
The PGE2, PGE1, and PGD2 products formed in the pathways just cited can undergo a spontaneous dehydration reaction to form PGA2, PGA1, and PGJ2, respectively; PGJ2 may then undergo a spontaneous isomerization followed by a dehydration reaction to form in series Δ12-PGJ2 and 15-deoxy-Δ12,14-PGJ2.
PGH2 has a 5-carbon ring bridged by molecular oxygen. Its derived PGS have lost this oxygen bridge and contain a single, unsaturated 5-carbon ring with the exception of thromboxane A2 which possesses a 6-member ring consisting of one oxygen and 5 carbon atoms. The 5-carbon ring of prostacyclin is conjoined to a second ring consisting of 4 carbon and one oxygen atom. And, the 5 member ring of the cyclopentenone prostaglandins possesses an unsaturated bond in a conjugated system with a carbonyl group that causes these PGs to form bonds with a diverse range of bioactive proteins (for more see the diagrams at Prostanoid).
Hydroxyeicosatetraenoate (HETE) and leukotriene (LT) pathways
See Leukotriene#Biosynthesis, Hydroxyeicosatetraenoic acid, and Eoxin#Human biosynthesis.
The enzyme 5-lipoxygenase (5-LO or ALOX5) converts arachidonic acid into 5-hydroperoxyeicosatetraenoic acid (5-HPETE), which may be released and rapidly reduced to 5-hydroxyeicosatetraenoic acid (5-HETE) by ubiquitous cellular glutathione-dependent peroxidases. Alternately, ALOX5 uses its LTA synthase activity to act convert 5-HPETE to leukotriene A4 (LTA4). LTA4 is then metabolized either to LTB4 by Leukotriene A4 hydrolase or Leukotriene C4 (LTC4) by either LTC4 synthase or microsomal glutathione S-transferase 2 (MGST2). Either of the latter two enzymes act to attach the sulfur of cysteine's thio- (i.e. SH) group in the tripeptide glutamate-cysteine-glycine to carbon 6 of LTA4 thereby forming LTC4. After release from its parent cell, the glutamate and glycine residues of LTC4 are removed step-wise by gamma-glutamyltransferase and a dipeptidase to form sequentially LTD4 and LTE4. The decision to form LTB4 versus LTC4 depends on the relative content of LTA4 hydrolase versus LTC4 synthase (or glutathione S=transferase in cells; Eosinophils, mast cells, and alveolar macrophages possess relatively high levels of LTC4 synthase and accordingly form LTC4 rather than or to a far greater extent than LTB4. 5-LOX may also work in series with cytochrome P450 oxygenases or aspirin-treated COX2 to form Resolvins RvE1, RvE2, and 18S-RvE1 (see Specialized pro-resolving mediators#EPA-derived resolvins).
The enzyme arachidonate 12-lipoxygenase (12-LO or ALOX12) metabolizes arachidonic acid to the S stereoisomer of 12-hydroperoxyeicosatetraenoic acid (5-HPETE) which is rapidly reduced by cellular peroxidases to the S stereoisomer of 12-hydroxyeicosatetraenoic acid (12-HETE) or further metabolized to hepoxilins (Hx) such as HxA3 and HxB.
The enzymes 15-lipoxygenase-1 (15-LO-1 or ALOX15) and 15-lipoxygenase-2 (15-LO-2, ALOX15B) metabolize arachidonic acid to the S stereoisomer of 15-Hydroperoxyeicosatetraenoic acid (15(S)-HPETE) which is rapidly reduced by cellular peroxidases to the S stereoisomer of 15-Hydroxyicosatetraenoic acid (15(S)-HETE). The 15-lipoxygenases (particularly ALOX15) may also act in series with 5-lipoxygenase, 12-lipoxygenase, or aspirin-treated COX2 to form the lipoxins and epi-lipoxins or with P450 oxygenases or aspirin-treated COX2 to form Resolvin E3 (see Specialized pro-resolving mediators#EPA-derived resolvins.
A subset of Cytochrome P450 (CYP450) microsome-bound ω-hydroxylases (see 20-Hydroxyeicosatetraenoic acid) metabolize arachidonic acid to 20-Hydroxyeicosatetraenoic acid (20-HETE) and 19-hydroxyeicosatetraenoic acid by an omega oxidation reaction.
Epoxyeicosanoid pathway
The human cytochrome P450 (CYP) epoxygenases, CYP1A1, CYP1A2, CYP2C8, CYP2C9, CYP2C18, CYP2C19, CYP2E1, CYP2J2, and CYP2S1 metabolize arachidonic acid to the non-classic Epoxyeicosatrienoic acids (EETs) by coverting one of the fatty acid's double bonds to its epoxide to form one or more of the following EETs, 14,15-ETE, 11,12-EET, 8,9-ETE, and 4,5-ETE. 14,15-EET and 11,12-EET are the major EETs produced by mammalian, including human, tissues. The same CYPs but also CYP4A1, CYP4F8, and CYP4F12 metabolize eicosapentaenoic acid to five epoxide epoxyeicosatetraenoic acids (EEQs) viz., 17,18-EEQ, 14,15-EEQ, 11,12-EEQ. 8,9-EEQ, and 5,6-EEQ (see epoxyeicosatetraenoic acid).
Function, pharmacology, and clinical significance
The following table lists a sampling of the major eicosanoids that possess clinically relevant biological activity, the cellular receptors (see Cell surface receptor) that they stimulate or, where noted, antagonize to attain this activity, some of the major functions which they regulate (either promote or inhibit) in humans and mouse models, and some of their relevancies to human diseases.
Prostanoids
Many of the prostanoids are know to mediate local symptoms of inflammation: vasoconstriction or vasodilation, coagulation, pain, and fever. Inhibition of COX-1 and/or the inducible COX-2 isoforms, is the hallmark of NSAIDs (non-steroidal anti-inflammatory drugs), such as aspirin. Prostanoids also activate the PPARγ members of the steroid/thyroid family of nuclear hormone receptors, and directly influence gene transcription. Prostanoids have numerous other relevancies to clinical medicine as evidence by their use, the use of their more stable pharmacological analogs, of the use of their receptor antagonists as indicated in the following chart.
Cyclopentenone prostaglandins
PGA1, PGA2, PGJ2, Δ12-PGJ2, and 15-deox-Δ12,14-PGJ2 exhibit a wide range of anti-inflammatory and inflammation-resolving actions in diverse animal models. They therefore appear to function in a manner similar to Specialized pro-resolving mediators although one of their mechanisms of action, forming covalent bonds with key signaling proteins, differs from those of the specialized pro-resolving mediators.
HETEs and oxo-ETEs
As indicated in their individual Wikipedia pages, 5-hydroxyeicosatetraenoic acid (which, like 5-oxo-eicosatetraenoic acid, acts through the OXER1 receptor), 5-oxo-eicosatetraenoic acid, 12-Hydroxyeicosatetraenoic acid, 15-Hydroxyeicosatetraenoic acid, and 20-Hydroxyeicosatetraenoic acid show numerous activities in animal and human cells as well as in animal models that are related to, for example, inflammation, allergic reactions, cancer cell growth, blood flow to tissues, and/or blood pressure. However, their function and relevancy to human physiology and pathology have not as yet been shown.
leukotrienes
The three cysteinyl leukotrienes, LTC4, LTD4, and LTE4, are potent bronchoconstrictors, increasers of vascular permeability in postcapillary venules, and stimulators of mucus secretion that are released from the lung tissue of asthmatic subjects exposed to specific allergens. They play a pathophysiological role in diverse types of immediate hypersensitivity reactions. Drugs that block their activation of the CYSLTR1 receptor viz., montelukast, zafirlukast, and pranlukast, are used clinically as maintenance treatment for allergen-induced asthma and rhinitis; nonsteroidal anti-inflammatory drug-induced asthma and rhinitis (see Aspirin-induced asthma); exercise- and cold-air induced asthma (see Exercise-induced bronchoconstriction); and childhood sleep apnea due to adenotonsillar hypertrophy (see Acquired non-inflammatory myopathy#Diet and Trauma Induced Myopathy). When combined with antihistamine drug therapy, they also appear useful for treating urticarial diseases such as hives.
Lipoxins and epi-lipoxins
LxA4, LxB4, 15-epi-LxA4, and 15-epi-LXB4, like other members of the specialized pro-resolving mediators) class of eicosanoids, possess anti-inflammatory and inflammation resolving activity. In a randomized controlled trial, AT-LXA4 and a comparatively stable analog of LXB4, 15R/S-methyl-LXB4, reduced the severity of eczema in a study of 60 infants and, in another study, inhaled LXA4 decreased LTC4-initiated bronchoprovocation in patients with asthma.
Eoxins
The eoxins (EXC4, EXD4, EXE5) are newly described. They stimulate vascular permeability in an ex vivo human vascular endothelial model system, and in a small study of 32 volunteers EXC4 production by eosinophils isolated from severe and aspirin-intolerant asthmatics was greater than that from healthy volunteers and mild asthmatic patients; these findings have been suggested to indicate that the eoxins have pro-inflammatory actions and therefore potentially involved in various allergic reactions. Production of eoxins by Reed-Sternburg cells has also led to suggestion that they are involve in Hodgkins disease. However, the clinical significance of eoxins has not yet been demonstrated.
Resolvin metabolites of eicosapentaenoic acid
RvE1, 18S-RvE1, RvE2, and RvE3, like other members of the specialized pro-resolving mediators) class of eicosanoids, possess anti-inflammatory and inflammation resolving activity. A synthetic analog of ReV1 is in clinical phase III testing (see Phases of clinical research) for the treatment of the inflammation-based dry eyesyndrome; along with this study, other clinical trials (NCT01639846, NCT01675570, NCT00799552 and NCT02329743) using an RvE1 analogue to treat various ocular conditions are underway. RvE1 is also in clinical development studies for the treatment of neurodegenerative diseases and hearing loss.
Other metabolites of eicosapentaenoic acid
The metabolites of eicosapentaenoic acid that are analogs of their arachidonic acid-derived prostanoid, HETE, and LT counterparts include: the 3-series prostanoids (e.g. PGE3, PGD3, PGF3α, PGI3, and TXA3), the hydroxyeicosapentaenoic acids (e.g. 5-HEPE, 12-HEPE, 15-HEPE, and 20-HEPE), and the 5-series LTs (e.g. LTB5, LTC5, LTD5, and LTE5). Many of the 3-series prostanoids, the hydroxyeicosapentaenoic acids, and the 5-series LT have been shown or though to be weaker stimulators of their target cells and tissues than their arachidonic acid-derived analogs. They are proposed to reduce the actions of their aracidonate-derived analogs by replacing their production with weaker analogs. Eicosapentaenoic acid-derived counterparts of the Eoxins have not been described.
Epoxyeicosanoids
The epoxy eicostrienoic acids (or EETs)—and, presumably, the epoxy eicosatetraenoic acids—have vasodilating actions on heart, kidney, and other blood vessels as well as on the kidney's reabsorption of sodium and water, and act to reduce blood pressure and ischemic and other injuries to the heart, brain, and other tissues; they may also act to reduce inflammation, promote the growth and metastasis of certain tumors, promote the growth of new blood vessels, in the central nervous system regulate the release of neuropeptide hormones, and in the peripheral nervous system inhibit or reduce pain perception.
The ω-3 and ω-6 series
Arachidonic acid (AA; 20:4 ω-6) sits at the head of the 'arachidonic acid cascade'—more than twenty different eicosanoid-mediated signaling paths controlling a wide array of cellular functions, especially those regulating inflammation, immunity, and the central nervous system.
In the inflammatory response, two other groups of dietary fatty acids form cascades that parallel and compete with the arachidonic acid cascade. EPA (20:5 ω-3) provides the most important competing cascade. DGLA (20:3 ω-6) provides a third, less prominent cascade. These two parallel cascades soften the inflammatory effects of AA and its products. Low dietary intake of these less-inflammatory fatty acids, especially the ω-3s, has been linked to several inflammation-related diseases, and perhaps some mental illnesses.
The U.S. National Institutes of Health and the National Library of Medicine state that there is 'A' level evidence that increased dietary ω-3 improves outcomes in hypertriglyceridemia, secondary cardiovascular disease prevention, and hypertension. There is 'B' level evidence ('good scientific evidence') for increased dietary ω-3 in primary prevention of cardiovascular disease, rheumatoid arthritis, and protection from ciclosporin toxicity in organ transplant patients. They also note more preliminary evidence showing that dietary ω-3 can ease symptoms in several psychiatric disorders.
Besides the influence on eicosanoids, dietary polyunsaturated fats modulate immune response through three other molecular mechanisms. They (a) alter membrane composition and function, including the composition of lipid rafts; (b) change cytokine biosynthesis; and (c) directly activate gene transcription. Of these, the action on eicosanoids is the best explored.
Mechanisms of ω-3 action
In general, the eicosanoids derived from AA promote inflammation, and those from EPA and from GLA (via DGLA) are less inflammatory, or inactive, or even anti-inflammatory and pro-resolving.
The figure shows the ω-3 and -6 synthesis chains, along with the major eicosanoids from AA, EPA, and DGLA.
Dietary ω-3 and GLA counter the inflammatory effects of AA's eicosanoids in three ways, along the eicosanoid pathways:
Role in inflammation
Since antiquity, the cardinal signs of inflammation have been known as: calor (warmth), dolor (pain), tumor (swelling), and rubor (redness). The eicosanoids are involved with each of these signs.
Redness—An insect's sting will trigger the classic inflammatory response. Short acting vasoconstrictors — TXA2—are released quickly after the injury. The site may momentarily turn pale. Then TXA2 mediates the release of the vasodilators PGE2 and LTB4. The blood vessels engorge and the injury reddens.
Swelling—LTB4 makes the blood vessels more permeable. Plasma leaks out into the connective tissues, and they swell. The process also loses pro-inflammatory cytokines.
Pain—The cytokines increase COX-2 activity. This elevates levels of PGE2, sensitizing pain neurons.
Heat—PGE2 is also a potent pyretic agent. Aspirin and NSAIDS—drugs that block the COX pathways and stop prostanoid synthesis—limit fever or the heat of localized inflammation.
History
In 1930, gynecologist Raphael Kurzrok and pharmacologist Charles Leib characterized prostaglandin as a component of semen. Between 1929 and 1932, Burr and Burr showed that restricting fat from animal's diets led to a deficiency disease, and first described the essential fatty acids. In 1935, von Euler identified prostaglandin. In 1964, Bergström and Samuelsson linked these observations when they showed that the "classical" eicosanoids were derived from arachidonic acid, which had earlier been considered to be one of the essential fatty acids. In 1971, Vane showed that aspirin and similar drugs inhibit prostaglandin synthesis. Von Euler received the Nobel Prize in medicine in 1970, which Samuelsson, Vane, and Bergström also received in 1982. E. J. Corey received it in chemistry in 1990 largely for his synthesis of prostaglandins.