
EC number 3.5.1.98 ExPASy NiceZyme view | CAS number 9076-57-7 | |
 | ||
Histone deacetylase hdac inhibitor cancer
Histone deacetylases (EC 3.5.1.98, HDAC) are a class of enzymes that remove acetyl groups (O=C-CH3) from an ε-N-acetyl lysine amino acid on a histone, allowing the histones to wrap the DNA more tightly. This is important because DNA is wrapped around histones, and DNA expression is regulated by acetylation and de-acetylation. Its action is opposite to that of histone acetyltransferase. HDAC proteins are now also called lysine deacetylases (KDAC), to describe their function rather than their target, which also includes non-histone proteins.
Contents
- Histone deacetylase hdac inhibitor cancer
- Histone deacetylases and histone deacetylase inhibitors video abstract 29965
- HDAC super family
- Classes of HDACs in higher eukaryotes
- Subtypes
- Subcellular distribution
- Histone modification
- Non histone effects
- HDAC inhibitors
- References
Histone deacetylases and histone deacetylase inhibitors video abstract 29965
HDAC super family
Together with the acetylpolyamine amidohydrolases and the acetoin utilization proteins, the histone deacetylases form an ancient protein superfamily known as the histone deacetylase superfamily.
Classes of HDACs in higher eukaryotes
HDACs, are classified in four classes depending on sequence homology to the yeast original enzymes and domain organization:
HDAC (except class III) contain zinc and are known as Zn-dependent histone deacetylases.
Subtypes
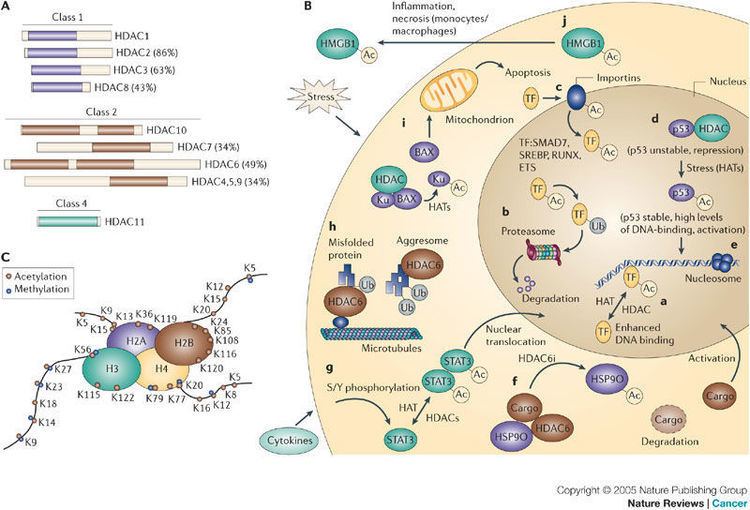
HDAC proteins are grouped into four classes (see above) based on function and DNA sequence similarity. Class I, II and IV are considered "classical" HDACs whose activities are inhibited by trichostatin A (TSA) and have a zinc dependent active site, whereas Class III enzymes are a family of NAD+-dependent proteins known as sirtuins and are not affected by TSA. Homologues to these three groups are found in yeast having the names: reduced potassium dependency 3 (Rpd3), which corresponds to Class I; histone deacetylase 1 (hda1), corresponding to Class II; and silent information regulator 2 (Sir2), corresponding to Class III. Class IV contains just one isoform (HDAC11), which is not highly homologous with either Rpd3 or hda1 yeast enzymes, and therefore HDAC11 is assigned to its own class. The Class III enzymes are considered a separate type of enzyme and have a different mechanism of action; these enzymes are NAD+-dependent, whereas HDACs in other classes require Zn2+ as a cofactor.
Subcellular distribution
Within the Class I HDACs, HDAC 1, 2, and 3 are found primarily in the nucleus, whereas HDAC8 is found in both the nucleus and the cytoplasm, and is also membrane-associated. Class II HDACs (HDAC4, 5, 6, 7 9, and 10) are able to shuttle in and out of the nucleus, depending on different signals.
HDAC6 is a cytoplasmic, microtuble-associated enzyme. HDAC6 deacetylates tubulin, Hsp90, and cortactin, and forms complexes with other partner proteins, and is, therefore, involved in a variety of biological processes.
Histone modification
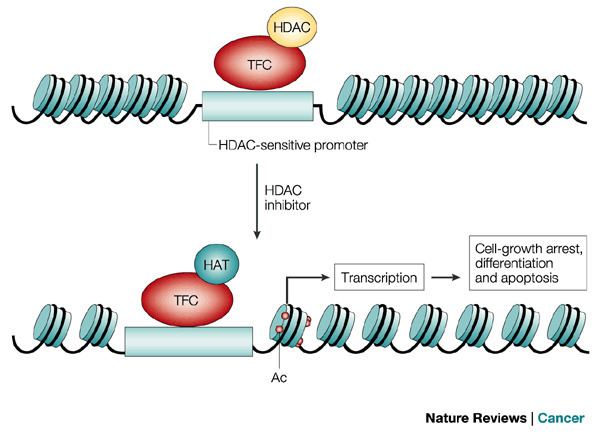
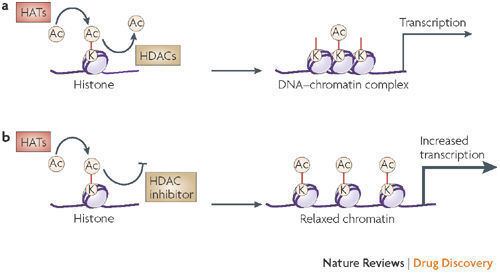
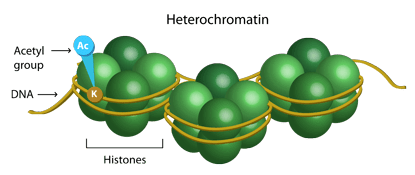
Histone tails are normally positively charged due to amine groups present on their lysine and arginine amino acids. These positive charges help the histone tails to interact with and bind to the negatively charged phosphate groups on the DNA backbone. Acetylation, which occurs normally in a cell, neutralizes the positive charges on the histone by changing amines into amides and decreases the ability of the histones to bind to DNA. This decreased binding allows chromatin expansion, permitting genetic transcription to take place. Histone deacetylases remove those acetyl groups, increasing the positive charge of histone tails and encouraging high-affinity binding between the histones and DNA backbone. The increased DNA binding condenses DNA structure, preventing transcription.
Histone deacetylase is involved in a series of pathways within the living system. According to the Kyoto Encyclopedia of Genes and Genomes (KEGG), these are:
Histone acetylation plays an important role in the regulation of gene expression. Hyperacetylated chromatin is transcriptionally active, and hypoacetylated chromatin is silent. A study on mice found that a specific subset of mouse genes (7%) was deregulated in the absence of HDAC1. Their study also found a regulatory crosstalk between HDAC1 and HDAC2 and suggest a novel function for HDAC1 as a transcriptional coactivator. HDAC1 expression was found to be increased in the prefrontal cortex of schizophrenia subjects, negatively correlating with the expression of GAD67 mRNA.
Non-histone effects
It is a mistake to regard HDACs solely in the context of regulating gene transcription by modifying histones and chromatin structure, although that appears to be the predominant function. The function, activity, and stability of proteins can be controlled by post-translational modifications. Protein phosphorylation is perhaps the most widely studied and understood modification in which certain amino acid residues are phosphorylated by the action of protein kinases or dephosphorylated by the action of phosphatases. The acetylation of lysine residues is emerging as an analogous mechanism, in which non-histone proteins are acted on by acetylases and deacetylases. It is in this context that HDACs are being found to interact with a variety of non-histone proteins—some of these are transcription factors and co-regulators, some are not. Note the following four examples:
These are just some examples of constantly emerging non-histone, non-chromatin roles for HDACs.
HDAC inhibitors
Histone deacetylase inhibitors (HDIs) have a long history of use in psychiatry and neurology as mood stabilizers and anti-epileptics, for example, valproic acid. In more recent times, HDIs are being studied as a mitigator or treatment for neurodegenerative diseases. Also in recent years, there has been an effort to develop HDIs for cancer therapy. Vorinostat (SAHA) was approved in 2006 for the treatment of cutaneous manifestations in patients with cutaneous T cell lymphoma (CTCL) that have failed previous treatments. A second HDI, Istodax (romidepsin), was approved in 2009 for patients with CTCL. The exact mechanisms by which the compounds may work are unclear, but epigenetic pathways are proposed. In addition, a clinical trial is studying valproic acid effects on the latent pools of HIV in infected persons. HDIs are currently being investigated as chemosensitizers for cytotoxic chemotherapy or radiation therapy, or in association with DNA methylation inhibitors based on in vitro synergy. Isoform selective HDIs which can aid in elucidating role of individual HDAC isoforms have been developed.
HDAC inhibitors have effects on non-histone proteins that are related to acetylation. HDIs can alter the degree of acetylation of these molecules and, therefore, increase or repress their activity. For the four examples given above (see Function) on HDACs acting on non-histone proteins, in each of those instances the HDAC inhibitor Trichostatin A (TSA) blocks the effect. HDIs have been shown to alter the activity of many transcription factors, including ACTR, cMyb, E2F1, EKLF, FEN 1, GATA, HNF-4, HSP90, Ku70, NFκB, PCNA, p53, RB, Runx, SF1 Sp3, STAT, TFIIE, TCF, YY1.
Histone deacetylase inhibitors may modulate the latency of some viruses, resulting in reactivation. This has been shown to occur, for instance, with a latent human herpesvirus-6 infection.