
Scientific name Pterosauria Extinction status Extinct | Higher classification Reptile | |
 | ||
Clade †PterosauromorphaPadian, 1997 Lower classifications | ||
Pterosaurs (/ˈTɛrəˌSɔːr, ˈtɛrOʊ-/; meaning "winged lizard") are flying reptiles of the extinct clade or order Pterosauria. They existed from the late Triassic to the end of the Cretaceous (228 to 66 million years ago). Pterosaurs are the earliest vertebrates known to have evolved powered flight. Their wings were formed by a membrane of skin, muscle, and other tissues stretching from the ankles to a dramatically lengthened fourth finger. Early species had long, fully toothed jaws and long tails, while later forms had a highly reduced tail, and some lacked teeth. Many sported furry coats made up of hair-like filaments known as pycnofibers, which covered their bodies and parts of their wings. Pterosaurs spanned a wide range of adult sizes, from the very small anurognathids to the largest known flying creatures of all time, including Quetzalcoatlus and Hatzegopteryx.
Contents
- What is a pterosaur
- Description
- Wings
- Parts of the wing
- Skull teeth and crests
- Pycnofibers
- History of discovery
- Flight
- Size
- Air sacs and respiration
- Nervous system
- Ground movement
- Diet and feeding habits
- Natural predators
- Reproduction and life history
- Daily activity patterns
- Origins
- Classification
- Extinction
- In popular culture
- References
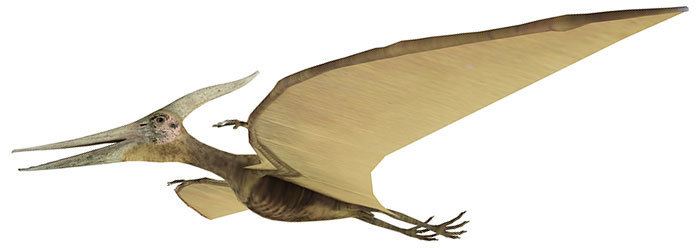
Pterosaurs are often referred to in the popular media and by the general public as "flying dinosaurs", but this is scientifically incorrect. The term "dinosaur" is restricted to just those reptiles descended from the last common ancestor of the groups Saurischia and Ornithischia (clade Dinosauria, which includes birds), and current scientific consensus is that this group excludes the pterosaurs, as well as the various groups of extinct marine reptiles, such as ichthyosaurs, plesiosaurs, and mosasaurs. Like the dinosaurs, and unlike these other reptiles, pterosaurs are more closely related to birds than to crocodiles or any other living reptile. Pterosaurs are also colloquially referred to as pterodactyls, particularly in fiction and by journalists. Technically, "Pterodactyl" refers only to members of the genus Pterodactylus, and more broadly to members of the suborder Pterodactyloidea of the pterosaurs.

What is a pterosaur
Description

The anatomy of pterosaurs was highly modified from their reptilian ancestors by the adaption to flight. Pterosaur bones were hollow and air-filled, like the bones of birds. They had a keeled breastbone that was developed for the attachment of flight muscles and an enlarged brain that shows specialised features associated with flight. In some later pterosaurs, the backbone over the shoulders fused into a structure known as a notarium, which served to stiffen the torso during flight, and provide a stable support for the scapula (shoulder blade).
Wings

Pterosaur wings were formed by membranes of skin and other tissues. The primary membranes attached to the extremely long fourth finger of each arm and extended along the sides of the body to the ankles.

While historically thought of as simple, leathery structures composed of skin, research has since shown that the wing membranes of pterosaurs were highly complex and dynamic structures suited to an active style of flight. The outer wings (from the tip to the elbow) were strengthened by closely spaced fibers called actinofibrils. The actinofibrils themselves consisted of three distinct layers in the wing, forming a crisscross pattern when superimposed on one another. The function of the actinofibrils is unknown, as is the exact material from which they were made. Depending on their exact composition (keratin, muscle, elastic structures, etc.), they may have been stiffening or strengthening agents in the outer part of the wing. The wing membranes also contained a thin layer of muscle, fibrous tissue, and a unique, complex circulatory system of looping blood vessels.

As evidenced by cavities in the wing bones of larger species and soft tissue preserved in at least one specimen, some pterosaurs extended their system of respiratory air sacs (see Paleobiology section below) into the wing membrane itself.
Parts of the wing
The pterosaur wing membrane is divided into three basic units. The first, called the propatagium ("first membrane"), was the forward-most part of the wing and attached between the wrist and shoulder, creating the "leading edge" during flight. This membrane may have incorporated the first three fingers of the hand, as evidenced in some specimens. The brachiopatagium ("arm membrane") was the primary component of the wing, stretching from the highly elongated fourth finger of the hand to the hind limbs (though where exactly on the hind limbs it anchored is controversial and may have varied between species, see below). Finally, at least some pterosaur groups had a membrane that stretched between the legs, possibly connecting to or incorporating the tail, called the uropatagium; the extent of this membrane is not certain, as studies on Sordes seem to suggest that it simply connected the legs but did not involve the tail (rendering it a cruropatagium). It is generally agreed though that non-pterodactyloid pterosaurs had a broader uro/cruropatagium, with pterodactyloids only having membranes running along the legs.
A bone unique to pterosaurs, known as the pteroid, connected to the wrist and helped to support a forward membrane (the propatagium) between the wrist and shoulder. Evidence of webbing between the three free fingers of the pterosaur forelimb suggests that this forward membrane may have been more extensive than the simple pteroid-to-shoulder connection traditionally depicted in life restorations. The position of the pteroid bone itself has been controversial. Some scientists, notably Matthew Wilkinson, have argued that the pteroid pointed forward, extending the forward membrane. This view was contradicted in a 2007 paper by Chris Bennett, who showed that the pteroid did not articulate as previously thought and could not have pointed forward, but rather inward toward the body as traditionally thought. Peters (2009) proposed that the pteroid articulated with the ‘saddle' of the radiale (proximal syncarpal) and both the pteroid and preaxial carpal were migrated centralia. This view of the articulation of the pteroid has since been supported by specimens of Changchengopterus pani and Darwinopterus linglongtaensis, both of which show the pteroid in articulation with the proximal syncarpal.
The pterosaur wrist consists of two inner (proximal) and four outer (distal) carpals (wrist bones), excluding the pteroid bone, which may itself be a modified distal carpal. The proximal carpals are fused together into a "syncarpal" in mature specimens, while three of the distal carpals fuse to form a distal syncarpal. The remaining distal carpal, referred to here as the medial carpal, but which has also been termed the distal lateral, or pre-axial carpal, articulates on a vertically elongate biconvex facet on the anterior surface of the distal syncarpal. The medial carpal bears a deep concave fovea that opens anteriorly, ventrally and somewhat medially, within which the pteroid articulates. In derived pterodactyloids like pteranodontians and azhdarchoids, metacarpals I-III are small and do not connect to the carpus, instead hanging in contact with the fourth metacarpal; in nyctosaurids the forelimb digits besides the wingfinger have been lost altogether.
There has been considerable argument among paleontologists about whether the main wing membranes (brachiopatagia) attached to the hind limbs, and if so, where. fossils of the rhamphorhynchoid Sordes, the anurognathid Jeholopterus, and a pterodactyloid from the Santana Formation seem to demonstrate that the wing membrane did attach to the hindlimbs, at least in some species. However, modern bats and flying squirrels show considerable variation in the extent of their wing membranes and it is possible that, like these groups, different species of pterosaur had different wing designs. Indeed, analysis of pterosaur limb proportions shows that there was considerable variation, possibly reflecting a variety of wing-plans.
Many, if not all, pterosaurs also had webbed feet.
Skull, teeth and crests
Most pterosaur skulls had elongated jaws with a full complement of needle-like teeth. In some cases, fossilized keratinous beak tissue has been preserved, though in toothed forms, the beak is small and restricted to the jaw tips and does not involve the teeth. Some advanced beaked forms were toothless, such as the pteranodonts and azhdarchids, and had larger, more extensive, and more bird-like beaks.
Unlike most archosaurs, the nasal and antorbital openings of pterodactyloid pterosaurs merged into a single large opening, called the nasoantorbital fenestra. This feature likely evolved to lighten the skull for flight.
Some species of pterosaurs featured elaborate crests. The first and perhaps best known of these is the distinctive backward-pointing crest of some Pteranodon species, though a few pterosaurs, such as the tapejarids and Nyctosaurus, sported extremely large crests that often incorporated keratinous or other soft tissue extensions of the bony crest base.
Since the 1990s, new discoveries and more thorough study of old specimens have shown that crests are far more widespread among pterosaurs than previously thought, due mainly to the fact that they were frequently extended by or composed completely of keratin, which does not fossilize as often as bone. In the case of pterosaurs like Pterorhynchus and Pterodactylus, the true extent of these crests has only been uncovered using ultraviolet photography. The discovery of Pterorynchus and Austriadactylus, both crested "rhamphorhynchoids", showed that even primitive pterosaurs had crests (previously, crests were thought to be restricted to the more advanced pterodactyloids).
Pycnofibers
At least some pterosaurs had hair-like filaments known as pycnofibers on the head and body, similar to, but not homologous (sharing a common structure) with, mammalian hair. A fuzzy integument was first reported from a specimen of Scaphognathus crassirostris in 1831 by Goldfuss, and recent pterosaur finds and the technology for histological and ultraviolet examination of pterosaur specimens have provided incontrovertible proof: pterosaurs had pycnofiber coats. Pycnofibers were not true hair as seen in mammals, but a unique structure that developed a similar appearance. Although, in some cases, actinofibrils (internal structural fibers) in the wing membrane have been mistaken for pycnofibers or true hair, some fossils, such as those of Sordes pilosus (which translates as "hairy demon") and Jeholopterus ninchengensis, do show the unmistakable imprints of pycnofibers on the head and body, not unlike modern-day bats, another example of convergent evolution. The head-coats do not cover the pterosaur's large jaws in many of the specimens found so far.
Some (Czerkas and Ji, 2002) have speculated that pycnofibers were an antecedent of proto-feathers, but the available impressions of pterosaur integuments are not like the "quills" found on many of the bird-like maniraptoran specimens in the fossil record. Pterosaur pycnofibers were structured similarly to theropod proto-feathers. Pycnofibers were flexible, short filaments, "only 5-7mm in some specimens" and rather simple, "apparently lacking any internal detail aside from a central canal". Pterosaur "pelts" found "preserved in concentrated, dense mats of fibers, similar to those found surrounding fossilized mammals" suggest coats with a thickness comparable to many Mesozoic mammals, at least on the parts of the pterosaur covered in pycnofibers. The coat thickness, and surface area covered, definitely varied by pterosaur species.
The presence of pycnofibers (and the demands of flight) imply that pterosaurs were endothermic (warm-blooded). The absence of pycnofibers on pterosaur wings suggests that the coat did not have an aerodynamic function, lending support to the idea that pycnofibers evolved to aid pterosaur thermoregulation, as is common in warm-blooded animals, insulation being necessary to conserve the heat created by an endothermic metabolism.
Pterosaur "hair" was so obviously distinct from mammalian fur and other animal integuments, it required a new, separate name. The term "pycnofiber", meaning "dense filament", was first coined in a paper on the soft tissue impressions of Jeholopterus by palaeontologist Alexander W.A. Kellner and colleagues in 2009. Research into the genetic code of American alligator embryos could suggest that pycnofibres, crocodile scutes and avian feathers are developmentally homologous, based on the construction of their beta-keratin.
History of discovery
The first pterosaur fossil was described by the Italian naturalist Cosimo Alessandro Collini in 1784. Collini misinterpreted his specimen as a seagoing creature that used its long front limbs as paddles. A few scientists continued to support the aquatic interpretation even until 1830, when the German zoologist Johann Georg Wagler suggested that Pterodactylus used its wings as flippers. Georges Cuvier first suggested that pterosaurs were flying creatures in 1801, and coined the name "Ptero-dactyle" in 1809 for the specimen recovered in Germany. However, due to the standardization of scientific names, the official name for this genus became Pterodactylus, though the name "pterodactyl" continued to be popularly and incorrectly applied to all members of Pterosauria. Paleontologists now avoid using "pterodactyl" and prefer the term "pterosaur". They relegate the term "pterodactyl" specifically for members of the genus Pterodactylus or more broadly for members of the suborder Pterodactyloidea.
Flight
The mechanics of pterosaur flight are not completely understood or modeled at this time.
Katsufumi Sato, a Japanese scientist, did calculations using modern birds and concluded that it was impossible for a pterosaur to stay aloft. In the book Posture, Locomotion, and Paleoecology of Pterosaurs it is theorized that they were able to fly due to the oxygen-rich, dense atmosphere of the Late Cretaceous period. However, both Sato and the authors of Posture, Locomotion, and Paleoecology of Pterosaurs based their research on the now outdated theories of pterosaurs being seabird-like, and the size limit does not apply to terrestrial pterosaurs, such as azhdarchids and tapejarids. Furthermore, Darren Naish concluded that atmospheric differences between the present and the Mesozoic were not needed for the giant size of pterosaurs.
Another issue that has been difficult to understand is how they took off. Earlier suggestions were that pterosaurs were largely cold-blooded gliding animals, deriving warmth from the environment like modern lizards, rather than burning calories. In this case, it was unclear how the larger ones of enormous size, with an inefficient cold-blooded metabolism, could manage a bird-like takeoff strategy, using only the hind limbs to generate thrust for getting airborne. Later research shows them instead as being warm-blooded and having powerful flight muscles, and using the flight muscles for walking as quadrupeds. Mark Witton of the University of Portsmouth and Mike Habib of Johns Hopkins University suggested that pterosaurs used a vaulting mechanism to obtain flight. The tremendous power of their winged forelimbs would enable them to take off with ease. Once aloft, pterosaurs could reach speeds of up to 120 kilometres per hour (75 mph) and travel thousands of kilometres.
In 1985, the Smithsonian Institution commissioned aeronautical engineer Paul MacCready to build a half-scale working model of Quetzalcoatlus northropi. The replica was launched with a ground-based winch. It flew several times in 1986 and was filmed as part of the Smithsonian's IMAX film On the Wing. However, the model was not anatomically correct and embodied vertical and horizontal tail stabilizers that pterosaurs did not have. It also had a longer tail, changing the weight distribution.
Size
Pterosaurs had a wide range of sizes, with wingspans ranging from 250 mm (10 in) at their smallest, to 10–11 m (33–36 ft) at their largest.
Air sacs and respiration
A 2009 study showed that pterosaurs had a lung-air sac system and a precisely controlled skeletal breathing pump, which supports a flow-through pulmonary ventilation model in pterosaurs, analogous to that of birds. The presence of a subcutaneous air sac system in at least some pterodactyloids would have further reduced the density of the living animal.
Like modern crocodilians, pterosaurs appeared to have had a hepatic piston, seeing as their shoulder-pectoral girdles were too inflexible to move the sternum as in birds, and they possessed strong gastralia. Thus, their respiratory system had characteristics comparable to both modern archosaur clades.
Nervous system
An X-ray study of pterosaur brain cavities revealed that the animals (Rhamphorhynchus muensteri and Anhanguera santanae) had massive flocculi. The flocculus is a brain region that integrates signals from joints, muscles, skin and balance organs.
The pterosaurs' flocculi occupied 7.5% of the animals' total brain mass, more than in any other vertebrate. Birds have unusually large flocculi compared with other animals, but these only occupy between 1 and 2% of total brain mass.
The flocculus sends out neural signals that produce small, automatic movements in the eye muscles. These keep the image on an animal's retina steady. Pterosaurs may have had such a large flocculus because of their large wing size, which would mean that there was a great deal more sensory information to process. The low relative mass of the flocculi in birds is also a result of birds having a much larger brain overall; though this has been considered an indication that pterosaurs lived in a structurally simpler environment or had less complex behaviour compared to birds, and recent studies of crocodilians and other reptiles show that it is common for sauropsids to achieve high intelligence levels with small brains.
Studies on the endocast of Allkaruen show that brain evolution in pterodactyloids was a modular process.
Ground movement
Pterosaurs' hip sockets are oriented facing slightly upwards, and the head of the femur (thigh bone) is only moderately inward facing, suggesting that pterosaurs had an erect stance. It would have been possible to lift the thigh into a horizontal position during flight, as gliding lizards do.
There was considerable debate whether pterosaurs ambulated as quadrupeds or as bipeds. In the 1980s, paleontologist Kevin Padian suggested that smaller pterosaurs with longer hindlimbs, such as Dimorphodon, might have walked or even run bipedally, in addition to flying, like road runners. However, a large number of pterosaur trackways were later found with a distinctive four-toed hind foot and three-toed front foot; these are the unmistakable prints of pterosaurs walking on all fours.
Fossil footprints show that pterosaurs stood with the entire foot in contact with the ground (plantigrade), in a manner similar to many mammals like humans and bears. Footprints from azhdarchids and several unidentified species show that pterosaurs walked with an erect posture with their four limbs held almost vertically beneath the body, an energy-efficient stance used by most modern birds and mammals, rather than the sprawled limbs of modern reptiles. Indeed, erect-limbs may be omnipresent in pterosaurs.
Though traditionally depicted as ungainly and awkward when on the ground, the anatomy of some pterosaurs (particularly pterodactyloids) suggests that they were competent walkers and runners. Early pterosaurs have long been considered particularly cumbersome locomotors due to the presence of large cruropatagia, but they too appear to have been generally efficient on the ground.
The forelimb bones of azhdarchids and ornithocheirids were unusually long compared to other pterosaurs, and, in azhdarchids, the bones of the arm and hand (metacarpals) were particularly elongated. Furthermore, as a whole, azhdarchid front limbs were proportioned similarly to fast-running ungulate mammals. Their hind limbs, on the other hand, were not built for speed, but they were long compared with most pterosaurs, and allowed for a long stride length. While azhdarchid pterosaurs probably could not run, they would have been relatively fast and energy efficient.
The relative size of the hands and feet in pterosaurs (by comparison with modern animals such as birds) may indicate the type of lifestyle pterosaurs led on the ground. Azhdarchid pterosaurs had relatively small feet compared to their body size and leg length, with foot length only about 25%–30% the length of the lower leg. This suggests that azhdarchids were better adapted to walking on dry, relatively solid ground. Pteranodon had slightly larger feet (47% the length of the tibia), while filter-feeding pterosaurs like the ctenochasmatoids had very large feet (69% of tibial length in Pterodactylus, 84% in Pterodaustro), adapted to walking in soft muddy soil, similar to modern wading birds.
Though clearly forelimb-based launchers, basal pterosaurs have hindlimbs well adapted for hopping, suggesting a connection with archosaurs such as Scleromochlus.
Diet and feeding habits
Traditionally, almost all pterosaurs were seen as surface-feeding piscivores, a view that still dominates popular culture. In reality, however, the majority of pterosaurs are now thought to have been terrestrial carnivores or insectivores.
One of the few groups that were never thought to be piscivores are the anurognathids; these were instead thought to be nocturnal, aerial insectivores, a view still maintained today. With highly flexible joints on the wing finger, a broad, triangular wing shape, large eyes and short tail, these pterosaurs were analogous to some of today's insectivorous bats, being capable of high manoeuvrability at relatively low speeds.
Dimorphodon has been envisioned as a puffin analogue in the past, but its jaw structure and gait, combined with its poor flight capabilities, indicate that it was a terrestrial/semiarboreal feeder. It seems to have been a predator of small mammals and squamates, and possibly also preyed on large insects.
Campylognathoides is seen most commonly as a terrestrial predator of small vertebrates, due to its robust dentition capable of delivering fatal bites to small tetrapods. Another view is that it was an aerial carnivore, pursuing other pterosaurs or swooping on small terrestrial vertebrates. Evidence for the latter lifestyle comes from the highly robust, "gorilla-like" humerus and a high-aspect wing morphology, which is similar to that of falcons and would provide the speed, agility and raw power to pursue and lift prey.
Eudimorphodonts can be divided into two major categories-those with long, robust wings similar to Campylognathoides, and those with long, slender wings. Species in the former category, including Carniadactylus and Eudimorphodon itself, were highly aerial animals and fast, agile flyers. The former was almost certainly insectivorous due to its small size; Eudimorphodon has been found with fish remains in its stomach, but as most carnivores or omnivores will also eat fish, its clear diet is uncertain. Meanwhile, the slender-winged species, such as Austriadactylus and Caviramus, were likely terrestrial/semiarboreal in habits. The dentition of Austriadactylus seems to be well-suited to eating small tetrapods. Caviramus has high bite forces, indicating an adaptation towards hard foods. It should be noted that all eudimorphodonts possessed well-developed molariform teeth and could chew their food, with Austriadactylus and Eudimorphodon having a pair of these molariform teeth developed into enlarged fangs.
Rhamphorhynchids can be roughly classified into two categories. One is the longirostrine bauplan, with long, slender wings, needle-like dentition and long, thin jaws, represented by species akin to Rhamphorhynchus itself or Dorygnathus. These taxa were piscivores. The other group is the "robust-jawed" bauplan, represented by species such as Sericipterus, Scaphognathus and Harpactognathus, which have more robust jaws and teeth (which were ziphont in Sericipterus), and shorter, broader wings. These were either terrestrial/aerial predators of vertebrates or corvid-like generalists.
Wukongopterids like Darwinopterus were first seen as aerial predators; however, as they lack the robust jaw structure or powerful flying muscles of Campylognathoides or the "robust-jawed" rhamphorhynchids, they are now seen as arboreal or semiterrestrial insectivores. Darwinopterus robustidens, in particular, seems to have been a beetle specialist.
Among pterodactyloids a greater variation in diet is present. Pteranodontia contained many piscivorous taxa, such as ornithocheirids, boreopterids, pteranodontids and nyctosaurids. Some amount of niche partitioning seems to have been present: ornithocheirids and the later nyctosaurs were aerial dip-feeders like today's frigatebirds, while boreopterids were freshwater diving animals similar to cormorants or Platanista river dolphins, and pteranodonts were pelagic plunge-divers akin to boobies and gannets. Exceptionally among this group, istiodactylids were terrestrial carnivores, and extremely unusual among vertebrates in that they were likely primarily scavengers.
In contrast, Azhdarchoidea mostly contained terrestrial pterosaurs. Tapejarids are considered terrestrial omnivores, feeding heavily on vegetation, but possibly also on small insects and vertebrates.
Dsungaripterids are traditionally thought of as being specialist molluscivores, using their powerful jaws to crush the shells of molluscs and crustaceans. While this remains the dominant view of dsungaripterid paleobiology today, some have suggested that they were generalistic omnivores eating a variety of hard foods, as dsungaripterids are better adapted to terrestrial movement rather than wading and are found in inland deposits.
Thalassodromids were terrestrial carnivores. Thalassodromeus itself is notable in that, despite being named after a fishing method known as "skim-feeding", it (like any other pterosaur) was physically incapable of this behaviour and wasn't even piscivorous. Instead, it seems to have been an unusually predatory pterosaur, pursuing relatively large prey, including those too large to be swallowed whole. Support for this lifestyle comes from a similar jaw shape to that of phorusrhacids.
Azhdarchids are now well-known as being terrestrial predators akin to ground hornbills or some storks, eating any prey item they could swallow whole. Two major exceptions to this are Hatzegopteryx, which was a robustly built raptorial predator of relatively large prey, including medium-sized dinosaurs; and Alanqa, a specialist molluscivore.
Lonchodectids are known to have had bodily proportions similar to those of azhdarchoids, and were probably similarly terrestrial. They are noted for rather unusual dentitions, however, with the possible member Prejanopterus possessing a bizarrely curved upper jaw that might indicate some specialised lifestyle.
Natural predators
Pterosaurs are known to have been eaten by theropods. In the 1 July 2004 edition of Nature, paleontologist Eric Buffetaut discusses an Early Cretaceous fossil of three cervical vertebrae of a pterosaur with the broken tooth of a spinosaur, most likely Irritator, embedded in it. The vertebrae are known not to have been eaten and exposed to digestion, as the joints are still articulated.
Reproduction and life history
Very little is known about pterosaur reproduction, and pterosaur eggs are very rare. The first known pterosaur egg was found in the quarries of Liaoning, the same place that yielded feathered dinosaurs. The egg was squashed flat with no signs of cracking, so evidently the eggs had leathery shells, as in modern lizards. This was supported by the description of an additional pterosaur egg belonging to the genus Darwinopterus, described in 2011, which also had a leathery shell and, also like modern reptiles but unlike birds, was fairly small compared to the size of the mother. In 2014 five unflattened eggs from the species Hamipterus tianshanensis were found in an Early Cretaceous deposit in northwest China. Examination of the shells by scanning electron microscopy showed the presence of a thin calcareous eggshell layer with a membrane underneath. A study of pterosaur eggshell structure and chemistry published in 2007 indicated that it is likely pterosaurs buried their eggs, like modern crocodiles and turtles. Egg-burying would have been beneficial to the early evolution of pterosaurs, as it allows for more weight-reducing adaptations, but this method of reproduction would also have put limits on the variety of environments pterosaurs could live in, and may have disadvantaged them when they began to face ecological competition from birds.
A Darwinopterus specimen showcases that at least some pterosaurs had a pair of functional ovaries, as opposed to the single functional ovivary in birds, dismissing the reduction of functional ovaries as a requirement for powered flight.
Wing membranes preserved in pterosaur embryos are well developed, suggesting that pterosaurs were ready to fly soon after birth. Fossils of pterosaurs only a few days to a week old (called "flaplings") have been found, representing several pterosaur families, including pterodactylids, rhamphorhinchids, ctenochasmatids and azhdarchids. All preserve bones that show a relatively high degree of hardening (ossification) for their age, and wing proportions similar to adults. In fact, many pterosaur flaplings have been considered adults and placed in separate species in the past. Additionally, flaplings are normally found in the same sediments as adults and juveniles of the same species, such as the Pterodactylus and Rhamphorhynchus flaplings found in the Solnhofen limestone of Germany, and Pterodaustro flaplings from Brazil. All are found in deep aquatic environment far from shore.
It is not known whether pterosaurs practiced any form of parental care, but their ability to fly as soon as they emerged from the egg and the numerous flaplings found in environments far from nests and alongside adults has led most researchers, including Christopher Bennett and David Unwin, to conclude that the young were dependent on their parents for a relatively short period of time, during a period of rapid growth while the wings grew long enough to fly, and then left the nest to fend for themselves, possibly within days of hatching. Alternatively, they may have used stored yolk products for nourishment during their first few days of life, as in modern reptiles, rather than depend on parents for food.
Growth rates of pterosaurs once they hatched varied across different groups. In more primitive, long-tailed pterosaurs ("rhamphorhynchoids"), such as Rhamphorhynchus, the average growth rate during the first year of life was 130% to 173%, slightly faster than the growth rate of alligators. Growth in these species slowed after sexual maturity, and it would have taken more than three years for Rhamphorhynchus to attain maximum size. In contrast, the more advanced, large pterodactyloid pterosaurs, such as Pteranodon, grew to adult size within the first year of life. Additionally, pterodactyloids had determinate growth, meaning that the animals reached a fixed maximum adult size and stopped growing.
Daily activity patterns
Comparisons between the scleral rings of pterosaurs and modern birds and reptiles have been used to infer daily activity patterns of pterosaurs. The pterosaur genera Pterodactylus, Scaphognathus, and Tupuxuara have been inferred to be diurnal, Ctenochasma, Pterodaustro, and Rhamphorhynchus have been inferred to be nocturnal, and Tapejara has been inferred to be cathemeral, being active throughout the day for short intervals. As a result, the possibly fish-eating Ctenochasma and Rhamphorhynchus may have had similar activity patterns to modern nocturnal seabirds, and the filter-feeding Pterodaustro may have had similar activity patterns to modern anseriform birds that feed at night. The differences between activity patterns of the Solnhofen pterosaurs Ctenochasma, Rhamphorhynchus, Scaphognathus, and Pterodactylus may also indicate niche partitioning between these genera.
Origins
Because pterosaur anatomy has been so heavily modified for flight, and immediate transitional fossil predecessors have not so far been described, the ancestry of pterosaurs is not fully understood. Several hypotheses have been advanced, including links to the avemetatarsalian-like Scleromochlus, an ancestry among the basal archosauriforms, like Euparkeria, or among the protorosaurs.
Two researchers, Chris Bennett (1996) and David Peters (2000), have found pterosaurs to be protorosaurs or closely related to them. Peters used a technique called DGS, which involves applying the digital tracing features of photo editing software to images of pterosaur fossils. Bennett only recovered pterosaurs as close relatives of the protorosaurs after removing characteristics of the hind limb from his analysis, in an attempt to test the idea that these characters are the result of convergent evolution between pterosaurs and dinosaurs. However, subsequent analysis by Dave Hone and Michael Benton (2007) could not reproduce this result. Hone and Benton found pterosaurs to be closely related to dinosaurs even without hind limb characters. They also criticized previous studies by David Peters, raising questions about whether conclusions reached without access to the primary evidence, that is, pterosaur fossils, can be held to have the same weight as conclusions based strictly on first-hand interpretation. Hone and Benton concluded that, although more primitive pterosauromorphs are needed to clarify their relationships, pterosaurs are best considered archosaurs, and specifically ornithodirans, given current evidence. In Hone and Benton's analysis, pterosaurs are either the sister group of Scleromochlus or fall between it and Lagosuchus on the ornithodiran family tree. Sterling Nesbitt (2011) found strong support for a clade composed of Scleromochlus and pterosaurs.
More recent studies on basal pterosaur hindlimb morphology seem to vindicate a connection to Scleromochlus. Like this archosaur, basal pterosaur lineages have plantigrade hindlimbs that show adaptations for saltitation.
Classification
In phylogenetic taxonomy, the clade Pterosauria has usually been defined as node-based and anchored to several extensively studied taxa as well as those thought to be primitive. One 2003 study defined Pterosauria as "The most recent common ancestor of the Anurognathidae, Preondactylus and Quetzalcoatlus and all their descendants." However, these types of definition would inevitably leave any related species that are slightly more primitive out of the Pterosauria. To remedy this, a new definition was proposed that would anchor the name not to any particular species but to an anatomical feature, the presence of an enlarged fourth finger that supports a wing membrane. A broader clade, Pterosauromorpha, has been defined as all ornithodirans more closely related to pterosaurs than to dinosaurs.
The internal classification of pterosaurs has historically been difficult, because there were many gaps in the fossil record. Starting from the 21st century, new discoveries are now filling in these gaps and giving a better picture of the evolution of pterosaurs. Traditionally, they were organized into two suborders: the Rhamphorhynchoidea, a "primitive" group of long-tailed pterosaurs, and the Pterodactyloidea, "advanced" pterosaurs with short tails. However, this traditional division has been largely abandoned. Rhamphorhynchoidea is a paraphyletic (unnatural) group, since the pterodactyloids evolved directly from them and not from a common ancestor, so, with the increasing use of cladistics, it has fallen out of favor among most scientists.
The precise relationships between pterosaurs is still unsettled. Many studies of pterosaur relationships in the past have included limited data and were highly contradictory. However, newer studies using larger data sets are beginning to make things clearer. The cladogram (family tree) below follows a phylogenetic analysis presented by Andres & Myers in 2013.
Extinction
It was once thought that competition with early bird species might have resulted in the extinction of many of the pterosaurs. By the end of the Cretaceous, only large species of pterosaurs are known (but see below). The smaller species were thought to have become extinct, their niche filled by birds. However, pterosaur decline (if actually present) seems unrelated to bird diversity, as ecological overlap between the two groups appears to be minimal. At the end of the Cretaceous period, the Cretaceous–Paleogene extinction event, which wiped out all non-avian dinosaurs and most avian dinosaurs as well, and many other animals, seems also to have taken the pterosaurs.
In the early 2010s, several new pterosaur taxa were discovered dating to the Campanian/Maastrichtian, such as the ornithocheirids Piksi and "Ornithocheirus", possible pteranodontids and nyctosaurids, several tapejarids and the indeterminate non-azhdarchid Navajodactylus. This suggests that Late Cretaceous pterosaur faunas were far more diverse than previously thought, possibly not even having declined significantly from the early Cretaceous.
Small sized pterosaur species apparently were present in the Csehbánya Formation, indicating a higher diversity of Late Cretaceous pterosaurs than previously accounted for. The recent findings of a small cat-sized adult azhdarchid further indicate that small pterosaurs from the Late Cretaceous might actually have simply been rarely preserved in the fossil record, helped by the fact that there is a strong bias against terrestrial small sized vertebrates such as juvenile dinosaurs, and that their diversity might actually had been much larger than previously thought.
At least some non-pterodactylod pterosaurs survived into the Late Cretaceous, postulating a lazarus taxa situation for late Cretaceous pterosaur faunas.
In popular culture
Pterosaurs have been a staple of popular culture for as long as their cousins the dinosaurs, though they are usually not featured as prominently in films, literature or other art. Additionally, while the depiction of dinosaurs in popular media has changed radically in response to advances in paleontology, a mainly outdated picture of pterosaurs has persisted since the mid 20th century.
While the generic term "pterodactyl" is often used to describe these creatures, the animals depicted frequently represent either Pteranodon or Rhamphorhynchus, or a fictionalized hybrid of the two. Many children's toys and cartoons feature "pterodactyls" with Pteranodon-like crests and long, Rhamphorhynchus-like tails and teeth, a combination that never existed in nature. However, at least one type of pterosaur did have the Pteranodon-like crest and teeth—for example, the Ludodactylus, a name that means "toy finger" for its resemblance to old, inaccurate children's toys. Also, some depictions of pterosaurs incorrectly identify them as "birds", when in real life they were flying reptiles, and birds are actually descended from theropod dinosaurs.
Pterosaurs were used in fiction in Arthur Conan Doyle's 1912 novel The Lost World, and subsequent 1925 film adaptation. They have been used in a number of films and television programs since, including the 1933 film King Kong, and 1966's One Million Years B.C.. In the latter, animator Ray Harryhausen had to add inaccurate bat-like wing fingers to his stop motion models in order to keep the membranes from falling apart, though this particular error was common in art even before the film was made. Pterosaurs were mainly absent from notable film appearances until 2001, with Jurassic Park III. However, paleontologist Dave Hone has noted that, even after the 40 intervening years, the pterosaurs in this film had not been significantly updated to reflect modern research. Among the errors he noted as persisting from the 1960s to the 2000s, were teeth even in toothless species (the Jurassic Park III pterosaurs were intended to be Pteranodon, which translates as "toothless wing"), nesting behavior that was known to be inaccurate by 2001, and leathery wings, rather than the taut membranes of muscle fiber that was actually present and required for pterosaur flight.
The prehistoric Pokémon Aerodactyl is likely to have been inspired by pterosaurs.
In most media appearances, pterosaurs are depicted as aerial predators similar to birds of prey, grasping human victims with their taloned feet. No pterosaur species known so far possesses prehensile feet; all known pterosaurs have flat, plantigrade feet with no opposable toes, often poorly muscled and, in the case of pteranodontians, generally proportionally small. However, some pterosaurs might have had raptorial tendencies; Thalassodromeus possesses powerful jaws akin to those of phorusrhacids, and Hatzegopteryx's short neck and more powerful jaws have been interpreted as a speciation on larger prey.