
Pronunciation /ˌɒksᵻˈtoʊsɪn/ CAS Number 50-56-6 Molar mass 1,007.19 g/mol Protein binding 30% | PubChem CID 439302 CAS ID 50-56-6 | |
 | ||
Metabolism Biological half-life 1–6 min (IV)~2 h (intranasal) | ||
Oxytocin (Oxt; /ˌɒksᵻˈtoʊsɪn/, /ˌɒksi-/) is a human peptide hormone and neuropeptide. Oxytocin is also used as a medication to facilitate childbirth. Oxytocin is normally produced by the paraventricular nucleus of the hypothalamus and released by the posterior pituitary. It plays a role in social bonding, sexual reproduction in both sexes, and during and after childbirth. Oxytocin is released into the bloodstream as a hormone in response to stretching of the cervix and uterus during labor and with stimulation of the nipples from breastfeeding. This helps with birth, bonding with the baby, and milk production. Oxytocin was discovered by Henry Dale in 1906. Its molecular structure was discovered in 1952.
Contents
- Biochemistry
- Biosynthesis
- Neural sources
- Non neural sources
- Male
- Female
- Evolution
- Biological function
- Physiological
- Psychological
- Bonding
- Drugs
- Fear and anxiety
- Mood and depression
- Sex differences
- Social
- Chemistry
- History
- References
Biochemistry
Estrogen has been found to increase the secretion of oxytocin and to increase the expression of its receptor, the oxytocin receptor, in the brain. In women, a single dose of estradiol has been found to be sufficient to increase circulating oxytocin concentrations.
Biosynthesis
The oxytocin peptide is synthesized as an inactive precursor protein from the OXT gene. This precursor protein also includes the oxytocin carrier protein neurophysin I. The inactive precursor protein is progressively hydrolyzed into smaller fragments (one of which is neurophysin I) via a series of enzymes. The last hydrolysis that releases the active oxytocin nonapeptide is catalyzed by peptidylglycine alpha-amidating monooxygenase (PAM).
The activity of the PAM enzyme system is dependent upon vitamin C (ascorbate), which is a necessary vitamin cofactor. By chance, sodium ascorbate by itself was found to stimulate the production of oxytocin from ovarian tissue over a range of concentrations in a dose-dependent manner. Many of the same tissues (e.g. ovaries, testes, eyes, adrenals, placenta, thymus, pancreas) where PAM (and oxytocin by default) is found are also known to store higher concentrations of vitamin C.

Oxytocin is known to be metabolized by the oxytocinase, leucyl/cystinyl aminopeptidase. Other oxytocinases are also known to exist. Amastatin, bestatin (ubenimex), leupeptin, and puromycin have been found to inhibit the enzymatic degradation of oxytocin, though they also inhibit the degradation of various other peptides, such as vasopressin, met-enkephalin, and dynorphin A.
Neural sources

In the hypothalamus, oxytocin is made in magnocellular neurosecretory cells of the supraoptic and paraventricular nuclei, and is stored in Herring bodies at the axon terminals in the posterior pituitary. It is then released into the blood from the posterior lobe (neurohypophysis) of the pituitary gland. These axons (likely, but dendrites have not been ruled out) have collaterals that innervate neurons in the nucleus accumbens, a brain structure where oxytocin receptors are expressed. The endocrine effects of hormonal oxytocin and the cognitive or behavioral effects of oxytocin neuropeptides are thought to be coordinated through its common release through these collaterals. Oxytocin is also produced by some neurons in the paraventricular nucleus that project to other parts of the brain and to the spinal cord. Depending on the species, oxytocin receptor-expressing cells are located in other areas, including the amygdala and bed nucleus of the stria terminalis.
In the pituitary gland, oxytocin is packaged in large, dense-core vesicles, where it is bound to neurophysin I as shown in the inset of the figure; neurophysin is a large peptide fragment of the larger precursor protein molecule from which oxytocin is derived by enzymatic cleavage.
Secretion of oxytocin from the neurosecretory nerve endings is regulated by the electrical activity of the oxytocin cells in the hypothalamus. These cells generate action potentials that propagate down axons to the nerve endings in the pituitary; the endings contain large numbers of oxytocin-containing vesicles, which are released by exocytosis when the nerve terminals are depolarised.
Non-neural sources
Endogenous oxytocin concentrations in the brain have been found to be as much as 1000-fold higher than peripheral levels.
Outside the brain, oxytocin-containing cells have been identified in several diverse tissues, including in females in the corpus luteum and the placenta, in males in the testicles' interstitial cells of Leydig, the retina, the adrenal medulla, the thymus and the pancreas. The finding of significant amounts of this classically "neurohypophysial" hormone outside the central nervous system raises many questions regarding its possible importance in these different tissues.
Male
The Leydig cells in some species have been shown to possess the biosynthetic machinery to manufacture testicular oxytocin de novo, to be specific, in rats (which can synthesize vitamin C endogenously), and in guinea pigs, which, like humans, require an exogenous source of vitamin C (ascorbate) in their diets.
Female
Oxytocin is synthesized by corpora lutea of several species, including ruminants and primates. Along with estrogen, it is involved in inducing the endometrial synthesis of prostaglandin F2α to cause regression of the corpus luteum.
Evolution
Virtually all vertebrates have an oxytocin-like nonapeptide hormone that supports reproductive functions and a vasopressin-like nonapeptide hormone involved in water regulation. The two genes are usually located close to each other (less than 15,000 bases apart) on the same chromosome, and are transcribed in opposite directions (however, in fugu, the homologs are further apart and transcribed in the same direction).
The two genes are believed to result from a gene duplication event; the ancestral gene is estimated to be about 500 million years old and is found in cyclostomata (modern members of the Agnatha).
Biological function
Oxytocin has peripheral (hormonal) actions, and also has actions in the brain. Its actions are mediated by specific, oxytocin receptors. The oxytocin receptor is a G-protein-coupled receptor that requires magnesium and cholesterol. It belongs to the rhodopsin-type (class I) group of G-protein-coupled receptors.
Studies have looked at oxytocin's role in various behaviors, including orgasm, social recognition, pair bonding, anxiety, and maternal behaviors.
Physiological
The peripheral actions of oxytocin mainly reflect secretion from the pituitary gland. The behavioral effects of oxytocin are thought to reflect release from centrally projecting oxytocin neurons, different from those that project to the pituitary gland, or that are collaterals from them. Oxytocin receptors are expressed by neurons in many parts of the brain and spinal cord, including the amygdala, ventromedial hypothalamus, septum, nucleus accumbens, and brainstem.
Psychological
Bonding
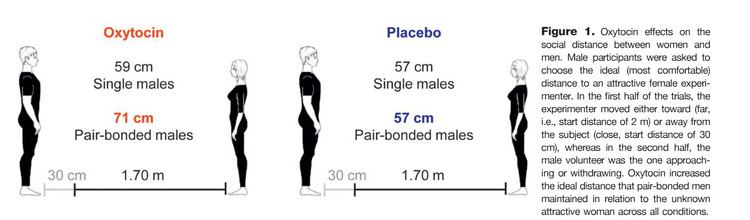
In the prairie vole, oxytocin released into the brain of the female during sexual activity is important for forming a monogamous pair bond with her sexual partner. Vasopressin appears to have a similar effect in males. Oxytocin has a role in social behaviors in many species, so it likely also does in humans. In a 2003 study, both humans and dog oxytocin levels in the blood rose after five to 24 minutes of a petting session. This possibly plays a role in the emotional bonding between humans and dogs.
Drugs
Fear and anxiety
Oxytocin is typically remembered for the effect it has on prosocial behaviors, such as its role in facilitating trust and attachment between individuals. Consequently, oxytocin is often referred to as the “love hormone". However, oxytocin has a more complex role than solely enhancing prosocial behaviors. There is consensus that oxytocin modulates fear and anxiety; that is, it does not directly elicit fear or anxiety. Two dominant theories explain the role of oxytocin in fear and anxiety. One theory states that oxytocin increases approach/avoidance to certain social stimuli and the second theory states that oxytocin increases the salience of certain social stimuli, causing the animal or human to pay closer attention to socially relevant stimuli.
Nasally administered oxytocin has been reported to reduce fear, possibly by inhibiting the amygdala (which is thought to be responsible for fear responses). Indeed, studies in rodents have shown oxytocin can efficiently inhibit fear responses by activating an inhibitory circuit within the amygdala. Some researchers have argued oxytocin has a general enhancing effect on all social emotions, since intranasal administration of oxytocin also increases envy and Schadenfreude. Individuals who receive an intranasal dose of oxytocin identify facial expressions of disgust faster than individuals who do not receive oxytocin. Facial expressions of disgust are evolutionarily linked to the idea of contagion. Thus, oxytocin increases the salience of cues that imply contamination, which leads to a faster response because these cues are especially relevant for survival. In another study, after administration of oxytocin, individuals displayed an enhanced ability to recognize expressions of fear compared to the individuals who received the placebo. Oxytocin modulates fear responses by enhancing the maintenance of social memories. Rats that are genetically modified to have a surplus of oxytocin receptors display a greater fear response to a previously conditioned stressor. Oxytocin enhances the aversive social memory, leading the rat to display a greater fear response when the aversive stimulus is encountered again.
Mood and depression
Oxytocin produces antidepressant-like effects in animal models of depression, and a deficit of it may be involved in the pathophysiology of depression in humans. The antidepressant-like effects of oxytocin are not blocked by a selective antagonist of the oxytocin receptor, suggesting that these effects are not mediated by the oxytocin receptor. In accordance, unlike oxytocin, the selective non-peptide oxytocin receptor agonist WAY-267,464 does not produce antidepressant-like effects, at least in the tail suspension test. In contrast to WAY-267,464, carbetocin, a close analogue of oxytocin and peptide oxytocin receptor agonist, notably does produce antidepressant-like effects in animals. As such, the antidepressant-like effects of oxytocin may be mediated by modulation of a different target, perhaps the vasopressin V1A receptor where oxytocin is known to weakly bind as an agonist.
Sildenafil has been found to enhance electrically evoked oxytocin release from the pituitary gland. In accordance, the drug shows oxytocin-dependent antidepressant-like effects in animals, and it has proposed that sildenafil may hold promise as a potential antidepressant in humans.
Sex differences
It has been shown that oxytocin differentially affects males and females. Females who are administered oxytocin are overall faster in responding to socially relevant stimuli than males who received oxytocin. Additionally, after the administration of oxytocin, females show increased amygdala activity in response to threatening scenes; however, males do not show increased amygdala activation. This phenomenon can be explained by looking at the role of gonadal hormones, specifically estrogen, which modulate the enhanced threat processing seen in females. Estrogen has been shown to stimulate the release of oxytocin from the hypothalamus and promote receptor binding in the amygdala.
It has also been shown that testosterone directly suppresses oxytocin in mice. This has been hypothesized to have evolutionary significance. With oxytocin suppressed, activities such as hunting and attacking invaders would be less mentally difficult as oxytocin is strongly associated with empathy.
Social
Chemistry
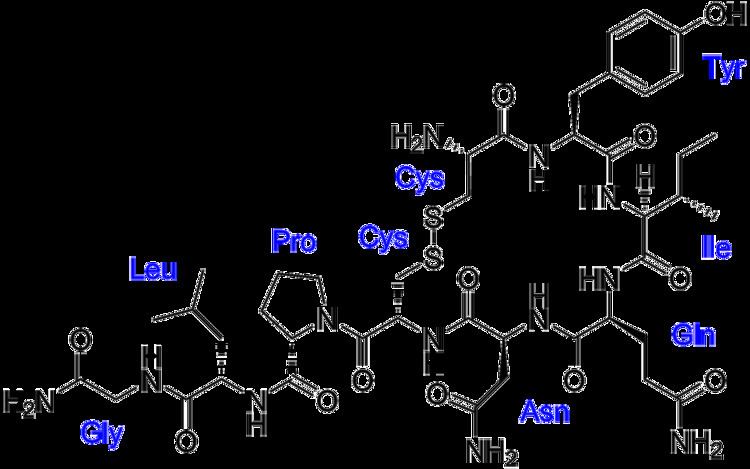
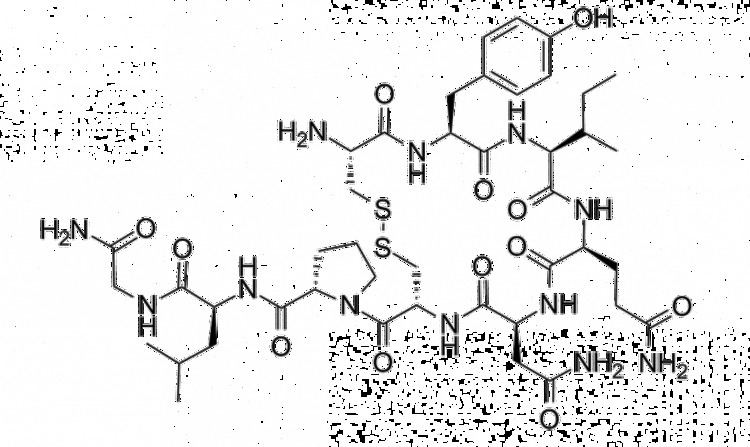
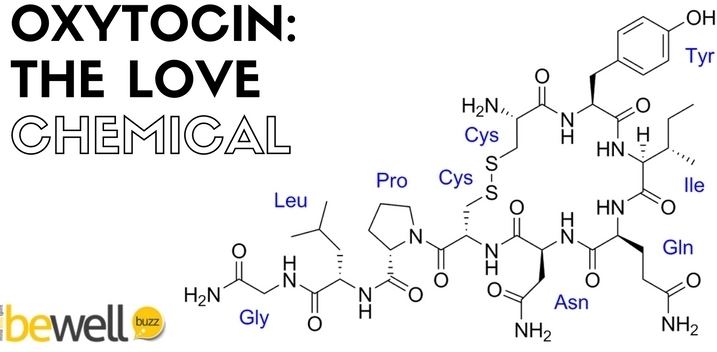
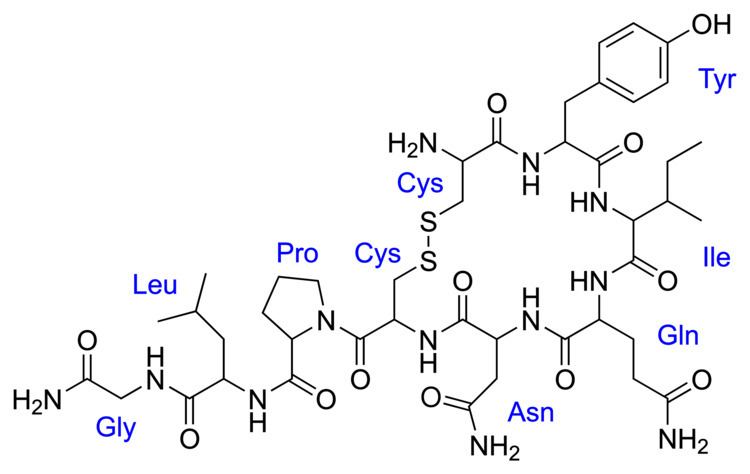
Oxytocin is a peptide of nine amino acids (a nonapeptide) in the sequence cysteine-tyrosine-isoleucine-glutamine-asparagine-cysteine-proline-leucine-glycine-amide (Cys – Tyr – Ile – Gln – Asn – Cys – Pro – Leu – Gly – NH2, or CYIQNCPLG-NH2); its C-terminus has been converted to a primary amide and a disulfide bridge joins the cysteine moieties. Oxytocin has a molecular mass of 1007 Da, and one international unit (IU) of oxytocin is the equivalent of about 2 μg of pure peptide.
While the structure of oxytocin is highly conserved in placental mammals, a novel structure of oxytocin was recently reported in marmosets, tamarins, and other new world primates. Genomic sequencing of the gene for oxytocin revealed a single in-frame mutation (thymine for cytosine) which results in a single amino acid substitution at the 8-position (proline for leucine). Since this original Lee et al. paper, two other laboratories have confirmed Pro8-OT and documented additional oxytocin structural variants in this primate taxon. Vargas-Pinilla et al. sequenced the coding regions of the OXT gene in other genera in new world primates and identified the following variants in addition to Leu8- and Pro8-OT: Ala8-OT, Thr8-OT, and Val3/Pro8-OT. Ren et al. identified a variant further, Phe2-OT in howler monkeys.
The biologically active form of oxytocin, commonly measured by RIA and/or HPLC techniques, is also known as the octapeptide "oxytocin disulfide" (oxidized form), but oxytocin also exists as a reduced straight-chain (non-cyclic) dithiol nonapeptide called oxytoceine. It has been theorized that oxytoceine may act as a free radical scavenger, as donating an electron to a free radical allows oxytoceine to be re-oxidized to oxytocin via the dehydroascorbate / ascorbate redox couple.
The structure of oxytocin is very similar to that of vasopressin. Both are nonapeptides with a single disulfide bridge, differing only by two substitutions in the amino acid sequence (differences from oxytocin bolded for clarity): Cys – Tyr – Phe – Gln – Asn – Cys – Pro – Arg – Gly – NH2. A table showing the sequences of members of the vasopressin/oxytocin superfamily and the species expressing them is present in the vasopressin article. Oxytocin and vasopressin were isolated and their total synthesis reported in 1954, work for which Vincent du Vigneaud was awarded the 1955 Nobel Prize in Chemistry with the citation: "for his work on biochemically important sulphur compounds, especially for the first synthesis of a polypeptide hormone."
Oxytocin and vasopressin are the only known hormones released by the human posterior pituitary gland to act at a distance. However, oxytocin neurons make other peptides, including corticotropin-releasing hormone and dynorphin, for example, that act locally. The magnocellular neurosecretory cells that make oxytocin are adjacent to magnocellular neurosecretory cells that make vasopressin. These are large neuroendocrine neurons which are excitable and can generate action potentials.
History
The word oxytocin was coined from the term oxytocic. Greek ὀξύς, oxys, and τόκος, tokos, meaning "quick birth")
Its uterine-contracting properties were discovered by British pharmacologist Sir Henry Hallett Dale in 1906. And its milk ejection property was described by Ott and Scott in 1910 and by Schafer and Mackenzie in 1911.
Oxytocin became the first polypeptide hormone to be sequenced or synthesized. Du Vigneaud was awarded the Nobel Prize in 1955 for his work.