
Subclass Elasmobranchii | Subphylum Vertebrata Superorder Selachimorpha Higher classification Carcharodon | |
 | ||
Genus Carcharodon or †Carcharocles Similar | ||
Megalodon (/ˈmɛɡəlOʊˌDɒN/ MEG-ə-lə-don, /ˈmeɪɡə-/ MAY-gə-; meaning "big tooth", from Ancient Greek: μέγας (megas) "big, mighty" and ὀδoύς (odoúS), "tooth"—whose stem is odont-, as seen in the genitive case form ὀδόντος, odóntos) is an extinct species of shark that lived approximately 23 to 2.6 million years ago, during the Cenozoic Era (Early Miocene to end of Pliocene).
Contents
- largest shark in the world megalodon hindi
- Glossopetrae
- Identification
- Fossils
- Taxonomy and evolution
- C megalodon within Carcharodon
- C megalodon within Carcharocles
- Reconsideration of megatooth lineage from Carcharocles to Otodus
- Megalodon as a chronospecies
- Mako sharks as closest relatives of great white sharks
- Considerations
- Anatomy
- Size estimation
- Length
- Body mass estimates
- Dentition and jaw mechanics
- Bite force
- Functional parameters of teeth
- Skeletal anatomy
- Range and habitat
- Prey relationships
- Competition and impact on marine communities
- Feeding strategies
- Nursery areas
- Extinction
- Oceanic cooling and sea level drops
- Decline in food supply
- Competitive pressures
- Multiple causes
- In fiction
- References

The taxonomic assignment of C. megalodon has been debated for nearly a century, and is still under dispute. The two major interpretations are Carcharodon megalodon (under family Lamnidae) or Carcharocles megalodon (under the family Otodontidae). Consequently, the scientific name of this species is commonly abbreviated C. megalodon in the literature.

Regarded as one of the largest and most powerful predators in vertebrate history, C. megalodon probably had a profound impact on the structure of marine communities. Fossil remains suggest that this giant shark reached a length of 18 metres (59 ft), and also indicate that it had a cosmopolitan distribution. Scientists suggest that C. megalodon looked like a stockier version of the great white shark, Carcharodon carcharias.
The tooth of C. megalodon is the state fossil of North Carolina.
largest shark in the world megalodon hindi
Glossopetrae

According to Renaissance accounts, gigantic, triangular fossil teeth often found embedded in rocky formations were once believed to be the petrified tongues, or glossopetrae, of dragons and snakes. This interpretation was corrected in 1667 by Danish naturalist Nicolaus Steno, who recognized them as shark teeth, and famously produced a depiction of a shark's head bearing such teeth. He described his findings in the book The Head of a Shark Dissected, which also contained an illustration of a C. megalodon tooth.
Identification
Swiss naturalist Louis Agassiz gave the shark its initial scientific name, Carcharodon megalodon, in 1835, in his research work Recherches sur les poissons fossiles (Research on fossil fish), which he completed in 1843. C. megalodon teeth are morphologically similar to the teeth of the great white shark, and on the basis of this observation, Agassiz assigned C. megalodon to the genus Carcharodon. While the scientific name is C. megalodon, it is often informally dubbed the "megatooth shark", "giant white shark" or "monster shark".
Fossils
C. megalodon is represented in the fossil record primarily by teeth and vertebral centra. As with all sharks, C. megalodon's skeleton was formed of cartilage rather than bone; this means that most fossil specimens are poorly preserved. While the earliest C. megalodon remains were reported from late Oligocene strata, around 28 million years old, a more reliable date for the origin of the species is the early Miocene, about 23 million years ago. Although fossils are mostly absent in strata extending beyond the Tertiary boundary, they have been reported from subsequent Pleistocene strata. It is believed that C. megalodon became extinct around the end of the Pliocene, probably about 2.6 million years ago; reported post-Pliocene C. megalodon teeth are thought to be reworked fossils. C. megalodon had a cosmopolitan distribution; its fossils have been excavated from many parts of the world, including Europe, Africa and both North and South America, as well as Puerto Rico, Cuba, Jamaica, the Canary Islands, Australia, New Zealand, Japan, Malta, the Grenadines and India. C. megalodon teeth have been excavated from regions far away from continental lands, such as the Mariana Trench in the Pacific Ocean.
The most common fossils of C. megalodon are its teeth. Diagnostic characteristics include: triangular shape, robust structure, large size, fine serrations, and visible V-shaped neck. C. megalodon teeth can measure over 180 millimetres (7.1 in) in slant height or diagonal length, and are the largest of any known shark species.
Some fossil vertebrae have been found. The most notable example is a partially preserved vertebral column of a single specimen, excavated in the Antwerp basin, Belgium by M. Leriche in 1926. It comprises 150 vertebral centra, with the centra ranging from 55 millimetres (2.2 in) to 155 millimetres (6.1 in) in diameter. However, scientists have claimed that considerably larger vertebral centra can be expected. A partially preserved vertebral column of another C. megalodon specimen was excavated from Gram clay in Denmark by Bendix-Almgeen in 1983. This specimen comprises 20 vertebral centra, with the centra ranging from 100 millimetres (3.9 in) to 230 millimetres (9.1 in) in diameter.
Taxonomy and evolution
Even after decades of research and scrutiny, controversy over C. megalodon phylogeny persists. Several shark researchers (e.g. J. E. Randall, A. P. Klimley, D. G. Ainley, M. D. Gottfried, L. J. V. Compagno, S. C. Bowman, and R. W. Purdy) insist that C. megalodon is a close relative of the great white shark. However, others (e.g. D. S. Jordan, H. Hannibal, E. Casier, C. DeMuizon, T. J. DeVries, D. Ward, and H. Cappetta) cite convergent evolution as the reason for the dental similarity. Such Carcharocles advocates have gained noticeable support. However, the original taxonomic assignment still has wide acceptance.
C. megalodon within Carcharodon
The traditional view is that C. megalodon should be classified within the genus Carcharodon along with the great white shark. The main reasons cited for this phylogeny are: (1) an ontogenetic gradation, whereby the teeth shift from coarse serrations as a juvenile to fine serrations as an adult, the latter resembling C. megalodon's; (2) morphological similarity of teeth of young C. megalodon to those of C. carcharias; (3) a symmetrical second anterior tooth; (4) a large intermediate tooth that is inclined mesially; and (5) upper anterior teeth that have a chevron-shaped neck area on the lingual surface. Carcharodon supporters suggest that C. megalodon and C. carcharias share a common ancestor, Palaeocarcharodon orientalis.
C. megalodon within Carcharocles
Around 1923, the genus Carcharocles was proposed by D. S. Jordan and H. Hannibal, to classify the shark C. auriculatus. Later on, Carcharocles proponents assigned C. megalodon to Carcharocles. Carcharocles proponents also suggest that the direct ancestor of the sharks belonging to Carcharocles is an ancient giant shark called Otodus obliquus, which lived during the Paleocene and Eocene epochs. According to Carcharocles supporters, Otodus obliquus evolved into Otodus aksuaticus, which evolved into Carcharocles auriculatus, and then into Carcharocles angustidens, and then into Carcharocles chubutensis, and then into C. megalodon. Hence, the immediate ancestor of C. megalodon is C. chubutensis, because it serves as the missing link between C. augustidens and C. megalodon and it bridges the loss of the "lateral cusps" that characterize C. megalodon.
Reconsideration of megatooth lineage from Carcharocles to Otodus
Shark researchers are reconsidering the genus of the entire Carcharocles lineage back to Otodus. Evidence for megalodon being a member of Otodus was strengthened 2016 with the publication of the genus Megalolamna. In their study, Shimada et al. concluded that reclassifying all members of Carcharocles to Otodus is necessary to make Otodus monophyletic, and that Otodus is the sister clade of Megalolamna within the family Otodontidae. Within Otodus, O. obliquus is thought to be the sister to the megatoothed lineage.
Megalodon as a chronospecies
Shark researcher David Ward elaborated on the evolution of Carcharocles by implying that this lineage, stretching from the Paleocene to the Pliocene, is of a single giant shark which gradually changed through time, suggesting a case of chronospecies. This assessment may be credible.
Mako sharks as closest relatives of great white sharks
Carcharocles proponents point out that the great white shark is closely related to the ancient shark Isurus hastalis, the "broad tooth mako", rather than to C. megalodon. One reason cited by paleontologist Chuck Ciampaglio is that the dental morphometrics (variations and changes in the physical form of objects) of I. hastalis and C. carcharias are remarkably similar. Another reason cited is that C. megalodon teeth have much finer serrations than C. carcharias teeth. Further evidence linking the great white shark more closely to ancient mako sharks, rather than to C. megalodon, was provided in 2009 – the fossilized remains of a form of the great white shark about 4 million years old were excavated from southwestern Peru in 1988. These remains demonstrate a likely shared ancestor of modern mako and great white sharks.
Considerations
Ciampaglio asserted that dental similarities between C. megalodon and the great white are superficial with noticeable morphometric differences between them, and that these findings are sufficient to warrant a separate genus. However, some Carcharodon proponents (i.e., M. D. Gottfried, and R. E. Fordyce) provided more arguments for a close relationship between the megatooth and the great white. With respect to the recent controversy regarding fossil lamnid shark relationships, overall morphology – particularly the internal calcification patterns – of the great white shark vertebral centra have been compared to well-preserved fossil centra from the megatooth, including C. megalodon and C. angustidens. The morphological similarity of these comparisons supports a close relationship of the giant fossil megatooth species to extant whites.
Gottfried and Fordyce pointed out that some great white shark fossils are about 16 million years old and predate the transitional Pliocene fossils. In addition, the Oligocene C. megalodon records contradict the suggestion that C. chubutensis is the immediate ancestor of C. megalodon. These records also indicate that C. megalodon co-existed with C. angustidens.
Some paleontologists argue that the genus Otodus should be used for sharks within the Carcharocles lineage and that the genus Carcharocles should be discarded.
Several Carcharocles proponents (i.e. C. Pimiento, D. J. Ehret, B. J. MacFadden, and G. Hubbell) claim that both species belong to the order Lamniformes, and in the absence of living members of the family Otodontidae, the great white shark is the species most ecologically analogous to C. megalodon.
Anatomy
Among extant species, the great white shark is regarded as the best analogue to C. megalodon. The lack of well-preserved fossil C. megalodon skeletons led scientists to rely on the great white shark as the basis of its reconstruction and size estimation.
Size estimation
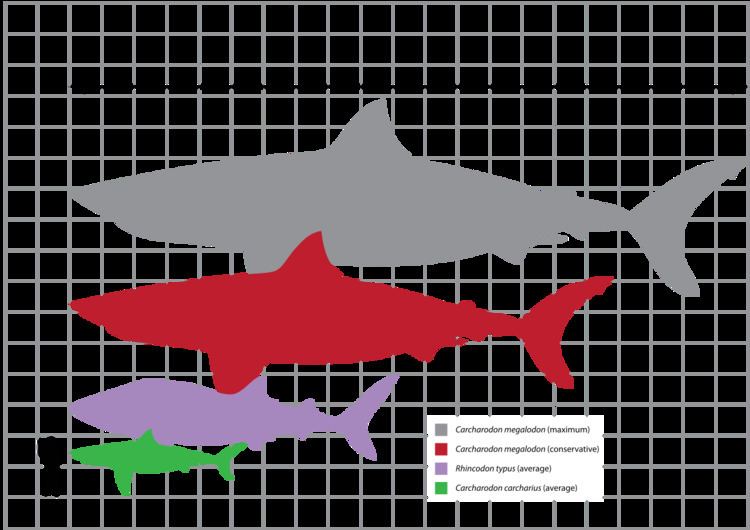
Due to fragmentary remains, estimating the size of C. megalodon has been challenging. However, the scientific community has concluded that C. megalodon was larger than the whale shark, Rhincodon typus. Scientists focused on two aspects of size: total length and body mass.
Length
The first attempt to reconstruct the jaw of C. megalodon was made by Bashford Dean in 1909. From the dimensions of this jaw reconstruction, it was hypothesized that C. megalodon could have approached 30 metres (98 ft). Better knowledge of dentition and more accurate muscle structures, led to a rectified version of Dean's jaw model about 70 percent of its original size and to a size consistent with modern findings. To resolve such errors, scientists, aided by new fossil discoveries of C. megalodon and improved knowledge of its closest living analogue's anatomy, introduced more quantitative methods for estimating its size based on the statistical relationships between the tooth sizes and body lengths. Some methods are mentioned below.
enamel height
In 1973, Hawaiian ichthyologist John E. Randall used a plotted graph to demonstrate a relationship between the enamel height (the vertical distance of the blade from the base of the enamel portion of the tooth to its tip) of the largest tooth in the upper jaw of the great white shark and the shark's total length. Randall extrapolated this method to estimate C. megalodon's total length. Randall cited two C. megalodon teeth in his work, specimen number 10356 at the American Museum of Natural History and specimen number 25730 at the United States National Museum, which had enamel heights of 115 millimetres (4.5 in) and 117.5 millimetres (4.63 in), respectively. These teeth yielded a corresponding total length of about 13 metres (43 ft). In 1991, Richard Ellis and John E. McCosker claimed that tooth enamel height does not necessarily increase in proportion to the animal's total length.
Largest anterior tooth height
In 1996, after scrutinizing 73 great white shark specimens, Michael D. Gottfried, Leonard Compagno and S. Curtis Bowman proposed a linear relationship between the shark's total length and the height of the largest upper anterior tooth. The proposed relationship is: total length in metres = − (0.096) × [UA maximum height (mm)]-(0.22). Gottfried and colleagues then extrapolated their technique to C. megalodon. The biggest C. megalodon tooth in the possession of this team, one discovered by Compagno in 1993, was an upper second anterior specimen, the maximum height of which was 168 millimetres (6.6 in). It yielded an estimated total length for C. megalodon of 15.9 metres (52 ft). Rumors of larger C. megalodon teeth persisted at the time. The maximum tooth height for this method is measured as a vertical line from the tip of the crown to the bottom of the lobes of the root, parallel to the long axis of the tooth. In layman's terms, the maximum height of the tooth is its slant height.
Root width
In 2002, shark researcher Clifford Jeremiah proposed that total length was proportional to the root width of an upper anterior tooth. He claimed that for every 1 centimetre (0.39 in) of root width, there are approximately 1.4 metres (4.6 ft) of shark length. Jeremiah pointed out that the jaw perimeter of a shark is directly proportional to its total length, with the width of the roots of the largest teeth being a tool for estimating jaw perimeter. The largest tooth in Jeremiah's possession had a root width of about 12 centimetres (4.7 in), which yielded 16.5 metres (54 ft) in total length. Ward asserted that this method is based on a sound principle that works well with most large sharks.
Crown height
In 2002, paleontologist Kenshu Shimada of DePaul University proposed a linear relationship between tooth crown height and total length in great white sharks after conducting anatomical analysis of several specimens. This relationship is expressed as: total length in centimetres = a + bx, where a is a constant, b is the slope of the line and x is the crown height of tooth in millimetres. This relationship allowed any tooth to be used for the estimate. The crown height was measured as maximum vertical enameloid height on the labial side. Shimada pointed out that previously proposed methods were based on weaker evaluation of dental homology, and that the growth rate between the crown and root is not isometric, which he considered in his model. Furthermore, this relationship could be used to predict the total length of sharks that are morphologically similar to the great white shark, such as C. megalodon. Using this model, the upper anterior tooth (with maximum height of 168 millimetres (6.6 in)) possessed by Gottfried and colleagues corresponded to a total length of 15.1 metres (50 ft). In 2010, shark researchers Catalina Pimiento, Dana J. Ehret, Bruce J. MacFadden and Gordon Hubbell estimated the total length of C. megalodon on the basis of Shimada's method. Among the specimens found in the Gatun Formation of Panama, specimen number 237956 yielded a total length of 16.8 metres (55 ft). Later on, shark researchers (including Pimiento, Ehret and MacFadden) revisited the Gatun Formation and recovered additional specimens. Specimen number 257579 yielded a total length of 17.9 metres (59 ft) on the basis of Shimada's method.
Consensus
In the 1990s, marine biologists such as Patrick J. Schembri and Staphon Papson opined that C. megalodon may have approached a maximum of around 24 to 25 metres (79 to 82 ft) in total length; however, Gottfried and colleagues asserted that C. megalodon could have reached a maximum of 20.3 metres (67 ft) in total length. However, a commonly acknowledged maximum total length of C. megalodon is 18 metres (59 ft).
Largest known specimens
Gordon Hubbell from Gainesville, Florida, possesses an upper anterior C. megalodon tooth whose maximum height is 184.1 millimetres (7.25 in). In addition, a C. megalodon jaw reconstruction contains a tooth whose maximum height is reportedly 193.67 millimetres (7.625 in). This jaw reconstruction was developed by fossil hunter Vito Bertucci, who was known as "Megalodon Man".
Body mass estimates
Gottfried and colleagues introduced a method to determine the mass of the great white after studying the length–mass relationship data of 175 specimens at various growth stages and extrapolated it to estimate C. megalodon's mass. According to their model, a 15.9 metres (52 ft) long C. megalodon would have a mass of about 48 metric tons (53 short tons), a 17 metres (56 ft) long C. megalodon would have a mass of about 59 metric tons (65 short tons), and a 20.3 metres (67 ft) long C. megalodon would have a mass of 103 metric tons (114 short tons).
Dentition and jaw mechanics
A team of Japanese scientists, T. Uyeno, O. Sakamoto, and H. Sekine, discovered and excavated partial remains of a C. megalodon, with its nearly complete associated set of teeth, from Saitama, Japan, in 1989. Another nearly complete associated C. megalodon dentition was excavated from the Yorktown Formations of Lee Creek, Aurora, North Carolina, in the United States and served as the basis of a jaw reconstruction of C. megalodon at the American Museum of Natural History in New York City. These associated tooth sets solved the mystery of how many teeth would be in each row of the jaws of C. megalodon. As a result, highly accurate jaw reconstructions became possible. More associated C. megalodon dentitions were found in later years. Based on these discoveries, scientists S. Applegate and L. Espinosa published an artificial dental formula (representation of dentition of an animal with respect to types of teeth and their arrangement within the animal's jaw) for C. megalodon in 1996. Most accurate modern C. megalodon jaw reconstructions are based on this dental formula.
The dental formula of C. megalodon is: 2.1.7.43.0.8.4.
As evident from the formula, C. megalodon had four kinds of teeth in its jaws.
C. megalodon had a very robust dentition, and had a total of about 276 teeth in its jaws, spanning 5 rows. Paleontologists suggest that a very large C. megalodon had jaws over 2 metres (6.6 ft) across.
Bite force
In 2008, a team of scientists led by S. Wroe conducted an experiment to determine the bite force of the great white shark, using a 2.5 metres (8.2 ft) long specimen, and then isometrically scaling the results for its maximum confirmed size and the conservative minimum and maximum body mass of C. megalodon, placing the bite force of the latter between 108,514 N (24,390 lbf) and 182,201 N (40,960 lbf) in a posterior bite. Compared to 18,216 N (4,095 lbf) for the largest confirmed great white shark, and 7,400 N (1,660 lbf) for the placoderm fish Dunkleosteus.
In addition, Wroe and colleagues pointed out that sharks shake sideways while feeding, amplifying the post-cranial generated forces. Therefore, the total force experienced by prey is probably higher than the estimate. The extraordinary bite forces in C. megalodon must be considered in the context of its great size and of paleontological evidence suggesting that C. megalodon was an active predator of large whales.
Functional parameters of teeth
The teeth of C. megalodon were exceptionally robust and serrated, which would have improved efficiency in slicing its prey's flesh. Paleontologist B. K. Kent suggested that these teeth are comparatively thicker for their size with much lower slenderness and bending strength ratios. Their roots are substantially larger relative to total tooth heights, and so have a greater mechanical advantage. Teeth with these traits are good cutting tools and are well suited for grasping powerful prey and would seldom crack even when slicing through bones.
Skeletal anatomy
Gottfried and colleagues further estimated the schematics of C. megalodon's entire skeleton. To support the beast's dentition, its jaws would have been massive, stouter, and more strongly developed than those of the great white, which possesses a comparatively gracile dentition. The jaws would have given it a "pig-eyed" profile. Its chondrocranium would have had a blockier and more robust appearance than that of the great white. Its fins were proportional to its larger size. Scrutiny of the partially preserved vertebral C. megalodon specimen from Belgium revealed that C. megalodon had a higher vertebral count than specimens of any known shark. Only the great white approached it.
Using the above characteristics, Gottfried and colleagues reconstructed the entire skeleton of C. megalodon, which was later put on display at the Calvert Marine Museum at Solomon's Island, Maryland, in the United States. This reconstruction is 11.5 metres (38 ft) long and represents a young individual. The team stresses that relative and proportional changes in the skeletal features of C. megalodon are ontogenetic in nature in comparison to those of the great white, as they occur in great white sharks while growing. Fossil remains of C. megalodon confirm that it had a heavily calcified skeleton while alive.
Range and habitat
Sharks, especially large species, are highly mobile and experience a complex life history amid wide distribution. Fossil records indicate that C. megalodon was cosmopolitan, and commonly occurred in subtropical to temperate latitudes. It has been found at latitudes up to 55° N; its inferred tolerated temperature range goes down to an annual mean of 12 °C (an annual range of 1–24 °C). It arguably had the capacity to endure such low temperatures by virtue of mesothermy, the physiological capability of large sharks to conserve metabolic heat by maintaining a higher body temperature than the surrounding water.
C. megalodon had enough adaptability to inhabit a wide range of marine environments (i.e., shallow coastal waters, areas of coastal upwelling, swampy coastal lagoons, sandy littorals, and offshore deep water environments), and exhibited a transient lifestyle. Adult C. megalodon were not abundant in shallow water environments, and mostly lurked offshore. C. megalodon may have moved between coastal and oceanic waters, particularly in different stages of its life cycle.
Fossil remains show a trend for specimens to be larger in the southern hemisphere than in the northern (mean lengths of 11.6 vs 9.6 m, respectively) and in the Pacific relative to the Atlantic (10.9 vs. 9.5 m). They do not, however, suggest any trend of changing body size with absolute latitude, or of change in size over time (although the megatooth lineage in general is thought to display a trend of increasing size over time). The overall modal length has been estimated at 10.5 m, with the length distribution skewed towards larger individuals, suggesting an ecological or competitive advantage for larger body size.
Prey relationships
Sharks generally are opportunistic predators, but scientists propose that C. megalodon was "arguably the most formidable carnivore ever to have existed". Its great size, high-speed swimming capability, and powerful jaws, coupled with a formidable killing apparatus, made it a super-predator capable of consuming a broad spectrum of fauna. A study about calcium isotopes of extinct and extant elasmobranchs revealed that C. megalodon fed at a higher trophic level than the contemporaneous great white shark.
Fossil evidence indicates that C. megalodon preyed upon cetaceans (i.e., dolphins), small whales (including cetotheriids such as Piscobalaena nana, squalodontids, and Odobenocetops), and large whales, (including sperm whales, bowhead whales, and rorquals), pinnipeds, porpoises, sirenians, and giant sea turtles. Marine mammals were regular prey targets for C. megalodon. Many whale bones have been found with clear signs of large bite marks (deep gashes) made by teeth that match the teeth of C. megalodon. Various excavations have revealed C. megalodon teeth lying close to the chewed remains of whales, and sometimes in direct association with them. Fossil evidence of interactions between C. megalodon and pinnipeds also exist. In one interesting observation, a 127 millimetres (5.0 in) C. megalodon tooth was found lying very close to a bitten earbone of a sea lion.
The feeding ecology of C. megalodon appears to have varied with age and between sites, like the modern great white. It is plausible that the adult C. megalodon population off the coast of Peru targeted primarily cetotheriid whales 2.5–7 metres (8 ft 2 in–23 ft 0 in) in length and other prey smaller than itself, rather than large whales in the same size class as themselves. Meanwhile, juveniles likely had a diet that consisted more of fish.
Competition and impact on marine communities
C. megalodon faced a highly competitive environment. However, its position at the top of the food chain probably had a profound impact on the structuring of marine communities. Fossil evidence indicates a correlation between C. megalodon emergence and extensive diversification of cetaceans. Juvenile C. megalodon preferred habitats where small cetaceans were abundant, and adult C. megalodon preferred habitats where large cetaceans were abundant. Such preferences may have developed shortly after they appeared in the Oligocene.
C. megalodon were contemporaneous with macro-predatory odontocetes (particularly raptorial sperm whales and squalodontids), which were also probably among the era's apex predators, and provided competition. In response to competition from giant macro-predatory sharks, macro-predatory odontocetes may have evolved defensive adaptations; some species became pack predators, and some attained gigantic sizes, such as Livyatan melvillei. By Late Miocene, raptorial sperm whales experienced a significant decline in abundance and diversity. However, raptorial delphinids began to emerge during the Pliocene, to fill this ecological void.
Like other sharks, C. megalodon also would have been piscivorous. Fossil evidence indicates that other notable species of macro-predatory sharks (e.g., great white sharks) responded to competitive pressure from C. megalodon by avoiding regions it inhabited. C. megalodon probably also had a tendency for cannibalism.
Feeding strategies
Sharks often employ complex hunting strategies to engage large prey animals. Some paleontologists suggest that great white shark hunting strategies may offer clues as to how C. megalodon hunted its unusually large prey. However, fossil evidence suggests that C. megalodon employed even more effective hunting strategies against large prey than the great white shark.
Paleontologists surveyed fossils to determine attacking patterns. One particular specimen – the remains of a 9 metres (30 ft) long prehistoric baleen whale (of an unknown Miocene taxon) – provided the first opportunity to quantitatively analyze its attack behavior. The predator primarily focused on the tough bony portions (i.e., shoulders, flippers, rib cage, and upper spine) of the prey, which great white sharks generally avoid. Dr. B. Kent elaborated that C. megalodon attempted to crush the bones and damage delicate organs (i.e., heart and lungs) harbored within the rib cage. Such an attack would have immobilized the prey, which would have died quickly from injuries to these vital organs. These findings also clarify why the ancient shark needed more robust dentition than that of the great white shark. Furthermore, attack patterns could differ for prey of different sizes. Fossil remains of some small cetaceans (e.g. cetotheriids) suggest that they were rammed with great force from below before being killed and eaten.
During the Pliocene, larger and more advanced cetaceans appeared. C. megalodon apparently further refined its hunting strategies to cope with these large whales. Numerous fossilized flipper bones (i.e., segments of the pectoral fins) and caudal vertebrae of large whales from the Pliocene have been found with C. megalodon bite marks. This paleontological evidence suggests that C. megalodon would immobilize a large whale by ripping apart or biting off its locomotive structures before killing and feeding on it.
Nursery areas
Fossil evidence suggests that the preferred nursery sites of C. megalodon were warm water coastal environments, where threats were minor and food plentiful. Nursery sites were identified in the Gatun Formation of Panama, the Calvert Formation of Maryland, Banco de Concepción in the Canary Islands, and the Bone Valley Formation of Florida. Given that all extant lamniform sharks give birth to live young, this is believed to have been true of C. megalodon also. The size of neonate C. megalodon teeth indicate that pups were around 2 to 4 metres (6.6 to 13.1 ft) in total length at birth. Their dietary preferences display an ontogenetic shift. Young C. megalodon commonly preyed on fish, giant sea turtles, dugongs and small cetaceans; mature C. megalodon moved to off-shore cetacean high-use areas and consumed large cetaceans.
However, an exceptional case in the fossil record suggests that juvenile C. megalodon may occasionally have attacked much larger balaenopterid whales. Three tooth marks apparently from a 4–7-metre (13.1–23.0 ft) long Pliocene macro-predatory shark were found on a rib from an ancestral great blue or humpback whale that showed evidence of subsequent healing. Scientists suspect that this shark was a juvenile C. megalodon.
Extinction
The subject of C. megalodon’s extinction remains under investigation. Several possible causes for its decline and eventual disappearance have been proposed.
Oceanic cooling and sea level drops
The Earth has been in a long term cooling trend since the Miocene Climactic Optimum, 15–17 Ma ago. This trend may have been accelerated by changes in global ocean circulation caused by the closure of the Central American Seaway and/or other factors (see Pliocene climate), setting the stage for glaciation in the northern hemisphere. Consequently, during the late Pliocene and Pleistocene, there were ice ages, which cooled the oceans significantly. Expansion of glaciation during the Pliocene tied up huge volumes of water in continental ice sheets, resulting in significant sea level drops. It has been argued that this cooling trend adversely impacted C. megalodon, as it preferred warmer waters, causing it to decline in abundance until its ultimate extinction at the end of the Pliocene. Fossil evidence confirms the absence of C. megalodon in regions around the world where water temperatures had significantly declined during the Pliocene. Furthermore, these oceanographic changes may have restricted many of the suitable warm water nursery sites for C. megalodon, hindering reproduction. Nursery areas are pivotal for the survival of many shark species, in part because they protect juveniles from predation.
A recent study, however, failed to show a correlation between oceanic cooling in the Miocene-Pliocene and C. megalodon's decline (see below).
Decline in food supply
baleen whales attained their greatest diversity during the Miocene, with over 20 recognized genera in comparison to only six extant genera. Such diversity presented an ideal setting to support a gigantic macropredator such as C. megalodon. However, by the end of the Miocene many species of mysticetes had gone extinct; surviving species may have been faster swimmers and thus more elusive prey. Furthermore, after the closure of the Central American Seaway, additional extinctions occurred in the marine environment, and faunal redistribution took place; tropical great whales decreased in diversity and abundance. The extinction of C. megalodon correlates with the decline of many small mysticete lineages; it is possible that it was quite dependent on them as a food source. Whale migratory patterns during the Pliocene have been reconstructed from the fossil record, suggesting that most surviving species of whales showed a trend towards polar regions. The cooling of the oceans during the Pliocene might have restricted the access of C. megalodon to polar regions, depriving it of its food source of large whales. As a result of these developments, the food supply for C. megalodon in regions it inhabited during the Pliocene, primarily in low-to-mid latitudes, was no longer sufficient to sustain it worldwide. C. megalodon was adapted to a specialized lifestyle, and this lifestyle was disturbed by these developments. Paleontologist Albert Sanders suggests that C. megalodon was too large to sustain itself on the declining tropical food supply. The resulting shortage of food sources in the tropics during Plio-Pleistocene times may have fueled cannibalism by C. megalodon. Juveniles were at increased risk from attacks by adults during times of starvation.
Competitive pressures
Large raptorial delphinids (members of genus Orcinus) evolved during the Pliocene, and probably filled the ecological void left by the disappearance of raptorial sperm whales at the end of the Miocene. A minority view is that competition from ancestral killer whales may have contributed to the shark's decline (another source suggests more generally that "competition with large odontocetes" may have been a factor). Fossil records indicate that these delphinids commonly occurred at high latitudes during the Pliocene, indicating that they could cope with the increasingly prevalent cold water temperatures. They also occurred in the tropics (e.g., Orcinus sp. in South Africa).
Multiple causes
Expert consensus has pointed to factors such as a cooling trend in the oceans and a shortage of food sources during Plio-Pleistocene times having played a significant role in the demise of C. megalodon.
However, a recent analysis of the distribution, abundance and climatic range of C. megalodon over geologic time suggests that biotic factors, i.e. dwindling numbers of prey species combined with competition from new macro-predators (raptorial sperm whales, great white sharks and killer whales), were the primary drivers of its extinction. The distribution of C. megalodon during the Miocene and Pliocene did not correlate with warming and cooling trends; while the abundance and distribution of C. megalodon declined during the Pliocene, C. megalodon did show a capacity to inhabit anti-tropical latitudes. C. megalodon was found in locations with a mean temperature ranging from 12 to 27 °C (with a total range of from 1 to 33 °C), indicating that the global extent of suitable habitat for C. megalodon should not have been greatly affected by the temperature changes that occurred.
The extinction of C. megalodon set the stage for further changes in marine communities. Average body size of baleen whales increased significantly after its disappearance. Other apex predators gained from the loss of this formidable species, in some cases spreading to regions where C. megalodon became absent.
In fiction
C. megalodon has been portrayed in several works of fiction, including films and novels, and continues to hold its place among the most popular subjects for fiction involving sea monsters. Many of these works posit that at least a relict population of C. megalodon survived extinction and lurk in the vast depths of the ocean, and that individuals may manage to surface, either by human intervention or by natural means. Jim Shepard's story "Tedford and the Megalodon" is an example of this. Such beliefs are usually inspired by discoveries of a C. megalodon tooth, such as that made by members of HMS Challenger in 1872, which some believed to be only 10,000 years old.
Some works of fiction (such as Shark Attack 3: Megalodon and Steve Alten's Meg series) incorrectly depict C. megalodon as being a species over 70 million years old, and to have lived during the time of the dinosaurs. The writers of the movie Shark Attack 3: Megalodon depicted this assumption by including an altered copy of Great White Shark by shark researcher Richard Ellis. The copy shown in the film had several pages that do not exist in the book. The author sued the film's distributor, Lions Gate Entertainment, asking for a halt to the film's distribution along with $150,000 in damages. Steve Alten's Meg: A Novel of Deep Terror is probably best known for portraying this inaccuracy with its prologue and cover artwork depicting C. megalodon killing a tyrannosaur in the sea.
The Animal Planet fictional documentary, Mermaids: The Body Found, included an encounter 1.6 million years ago between a pod of mermaids and a C. megalodon. Later, in August 2013, the Discovery Channel opened its annual Shark Week series with another film for television, Megalodon: The Monster Shark Lives, a controversial docufiction about the creature that presented alleged evidence in order to suggest that C. megalodon was still alive. This program received criticism for being completely fictional; for example, all of the supposed "scientists" depicted were paid actors. In 2014 Discovery re-aired The Monster Shark Lives, along with a new one-hour program, Megalodon: The New Evidence, and an additional fictionalized program entitled Shark of Darkness: Wrath of Submarine, resulting in further backlash from media sources and the scientific community.