
Scientific name Placodermi | ||
 | ||
Similar Dunkleosteus, Acanthodii, Arthrodira | ||
Paleo profile dunkleosteus
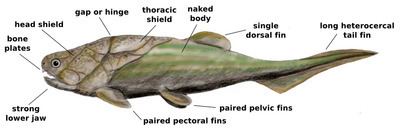
Placodermi (from the Greek πλάξ = plate and δέρμα = skin, literally "plate-skinned") is an extinct class of armoured prehistoric fish, known from fossils, which lived from the Silurian to the end of the Devonian Period. Their head and thorax were covered by articulated armoured plates and the rest of the body was scaled or naked, depending on the species. Placoderms were among the first jawed fish; their jaws likely evolved from the first of their gill arches. Placoderms are paraphyletic, and consist of several distinct outgroups or sister taxa to all living jawed vertebrates, which originated among their ranks. This is illustrated by a 419-million-year-old fossil, Entelognathus, from China, which is the only known placoderm with a type of bony jaw like that found in modern bony fishes. This includes a dentary bone, which is found in humans and other tetrapods,. The jaws in other placoderms were simplified and consisted of a single bone. Placoderms were also the first fish to develop pelvic fins, the precursor to hindlimbs in tetrapods, as well as true teeth. 380-million-year-old fossils of three other genera, Incistoscutum, Materpiscis and Austroptyctodus, represent the oldest known examples of live birth.
Contents
- Paleo profile dunkleosteus
- Characteristics
- Evolution and extinction
- Fossil record
- History of study
- Taxonomy and phylogeny
- Cladogram
- References

The first identifiable placoderms appear in the fossil record during the late Llandovery epoch of the early Silurian. The various groups of placoderms were diverse and abundant during the Devonian, but became extinct at the end-Devonian Hangenberg event 360 million years ago

Characteristics

Many placoderms, particularly the Rhenanida, Petalichthyida, Phyllolepida, and Antiarchi, were bottom-dwellers. In particular, the antiarchs, with their highly modified, jointed bony pectoral fins, were highly successful inhabitants of Middle-Late Devonian freshwater and shallow marine habitats, with the Middle to Late Devonian genus, Bothriolepis, known from over 100 valid species. The vast majority of placoderms were predators, many of which lived at or near the substrate. Many, primarily the Arthrodira, were active, nektonic predators that dwelled in the middle to upper portions of the water column. A study of the arthrodire Compagopiscis published in 2012 concluded that placoderms (at least this particular genus) likely possessed true teeth contrary to some early studies. The teeth had well defined pulp cavities and were made of both bone and dentine. However the tooth and jaw development were not as closely integrated as in modern gnathostomes. These teeth were likely homologous to the teeth of other gnathostomes.
One of the largest known arthrodires, Dunkleosteus terrelli , was 6 m (20 ft) long, and is presumed to have had a large distribution, as its remains have been found in Europe, and North America and possibly Morocco. Some paleontologists regard it as the world's first vertebrate "superpredator", preying upon other predators. Other, smaller arthrodires, such as Fallacosteus and Rolfosteus, both of the Gogo Formation of Western Australia, had streamlined, bullet-shaped head armor, strongly supporting the idea that many, if not most, arthrodires were active swimmers, rather than passive ambush-hunters whose armor practically anchored them to the sea floor. Some placoderms were herbivorous, such as the Middle to Late Devonian arthrodire Holonema, and some were planktivores, such as the gigantic, 8 m (26 ft) long arthrodire, Titanichthys.

Extraordinary evidence of internal fertilization in a placoderm was afforded by the discovery in the Gogo Formation, near Fitzroy Crossing, Kimberley, Western Australia, of a small female placoderm, about 25 cm (10 in) in length, which died in the process of giving birth to a 6 cm ( 2 1⁄2 in) offspring and was fossilized with the umbilical cord intact. The fossil, named Materpiscis attenboroughi (after scientist David Attenborough), had eggs which were fertilised internally, the mother providing nourishment to the embryo and giving birth to live young. With this discovery, the placoderm became the oldest vertebrate known to have given birth to live young ("viviparous"), pushing the date of first viviparity back some 200 million years earlier than had been previously known. Specimens of the arthrodire Incisoscutum ritchei, also from the Gogo Formation, have been found with embryos inside them indicating this group also had live bearing ability. The males reproduced by inserting a long clasper into the female. Elongated basipterygia are also found on the phyllolepid placoderms, such as Austrophyllolepis and Cowralepis, both from the Middle Devonian of Australia, suggesting that the basiptergia were used in copulation.

The placoderm claspers are not homologous with the claspers in cartilaginous fishes. The similarities between the structures has been revealed to be an example of convergent evolution. While the claspers in cartilaginous fishes are specialized parts of their paired pelvic fins that have been modified for copulation due to changes in the hox genes hoxd13, the origin of the mating organs in placoderms most likely relied on different sets of hox genes and were structures that developed further down the body as an extra and independent pair of appendages, but which during development turned into body parts used for reproduction only. Because they were not attached to the pelvic fins, as are the claspers in fish like sharks, they were much more flexible and could probably be rotated forward.
Evolution and extinction
It was thought for a time that placoderms became extinct due to competition from the first bony fish and early sharks, given a combination of the supposed inherent superiority of bony fish and the presumed sluggishness of placoderms. With more accurate summaries of prehistoric organisms it is now thought that they systematically died out as marine and freshwater ecologies suffered from the environmental catastrophes of the Devonian/Carboniferous extinction events
Fossil record
The earliest identifiable placoderm fossils are from China and date to the early Silurian. At that time, they were already differentiated into antiarchs and arthrodires as well as other, more primitive groups. Earlier fossils of basal Placodermi have not yet been discovered.
The Silurian fossil record of the placoderms is both literally and figuratively fragmented. Until the discovery of Silurolepis (and then, the discoveries of Entelognathus and Qilinyu), Silurian-aged placoderm specimens consisted of fragments. Some of them have been tentatively identified as antiarch or arthrodire due to histological similarities; and many of them have not yet been formally described or even named. The most commonly cited example of a Silurian placoderm, Wangolepis of Silurian China and possibly Vietnam, is known only from a few fragments that currently defy attempts to place them in any of the recognized placoderm orders. So far, only three officially described Silurian placoderms are known from more than scraps:
The first officially described and oldest Silurian placoderm is an antiarch, Shimenolepis, which is known from distinctively ornamented plates from the late Llandovery of Hunan, China. Shimenolepis plates are very similar to the early Devonian yunnanolepid Zhanjilepis, also known from distinctively ornamented plates.
Paleontologists and placoderm specialists suspect that the scarcity of placoderms in the Silurian fossil record is due to placoderms' living in environments unconducive to fossil preservation, rather than a genuine scarcity. This hypothesis helps to explain the placoderms' seemingly instantaneous appearance and diversity at the very beginning of the Devonian.
During the Devonian, the placoderms went on to inhabit and dominate almost all known aquatic ecosystems, both freshwater and saltwater. But this diversity ultimately suffered many casualties during the extinction event at the Frasnian–Famennian boundary, the Late Devonian extinctions. The remaining species then died out during the Devonian/Carboniferous extinction event; not a single placoderm species has been confirmed to have survived into the Carboniferous.
History of study
The earliest studies of placoderms were published by Louis Agassiz, in his five volumes on fossil fishes, 1833–1843. In those days, the placoderms were thought to be shelled jawless fish akin to ostracoderms. Some naturalists even suggested that they were shelled invertebrates or even turtle-like vertebrates.
In the late 1920s, Dr. Erik Stensiö, at the Swedish Museum of Natural History in Stockholm, established the details of placoderm anatomy and identified them as true jawed fishes related to sharks. He took fossil specimens with well-preserved skulls and ground them away, one tenth of a millimeter at a time. Between each grinding, he made an imprint in wax. Once the specimens had been completely ground away (and so destroyed), he made enlarged, three-dimensional models of the skulls to examine the anatomical details more thoroughly. Many other placoderm specialists thought that Stensiö was trying to shoehorn placoderms into a relationship with sharks; however, as more fossils were found, placoderms were accepted as a sister group of chondrichthyians.
Much later, the exquisitely preserved placoderm fossils from Gogo reef changed the picture again. They showed that placoderms shared anatomical features not only with chondrichthyians but with other gnathostome groups as well. For example, Gogo placoderms show separate bones for the nasal capsules as in gnathostomes; in both sharks and bony fish those bones are incorporated into the braincase.
Placoderms also share certain anatomical features only with the jawless osteostracans; because of this, the theory that placoderms are the sister group of chondrichthyians has been replaced by the theory that placoderms are a group of stem gnathostomes.
Taxonomy and phylogeny
Currently Placodermi are divided into eight recognized orders. There are two further controversial orders. One order is the monotypic Stensioellida, containing the enigmatic Stensioella, and the other order is the equally enigmatic Pseudopetalichthyida. These orders are considered to be basal or primitive groups within Placodermi, though their precise placement within the class remains unsure. Fossils of both are currently known only from the Hunsruck lagerstatten.
Cladogram
The following cladogram shows the interrelationships of placoderms according to Carr et al. (2009):