
Descendants R1a (M420), R1b (M343) | Ancestor R (R-M207) | |
 | ||
Defining mutations M173/P241/Page29, CTS916/M611/PF5859, CTS997/M612/PF6111, CTS1913/M654, CTS2565/M663, CTS2680, CTS2908/M666/PF6123, CTS3123/M670, CTS3321/M673, CTS4075/M682, CTS5611/M694, CTS7085/M716/Y481, CTS8116/M730, F93/M621/PF6114, F102/M625/PF6116, F132/M632, F211/Y290, F245/M659/Y477, FGC189/Y305, L875/M706/PF6131/YSC0000288, L1352/M785/YSC0000230, M306/PF6147/S1, M640/PF6118, M643, M689, M691/CTS4862/PF6042/YSC0001281, M710/PF6132/YSC0000192, M748/YSC0000207, M781, P225, P231, P233, P234, P236, P238/PF6115, P242/PF6113, P245/PF6117, P286/PF6136, P294/PF6112, PF6120 | ||
Haplogroup R1, or R-M173, is a Y-chromosome DNA haplogroup. A primary subclade of Haplogroup R (R-M207), it is defined by the SNP M173. The other primary subclade of Haplogroup R is Haplogroup R2 (R-M479).
Contents
Males carrying R-M173 in modern populations appear to comprise two subclades: R1a and R1b, which are found mainly in populations native to Eurasia (except East and Southeast Asia). R-M173 contains the majority of representatives of haplogroup R in the form of its subclades, R1a and R1b (Rosser 2000, Semino 2000, and Genographic 2011).
Karafet et al. (2014) "rapid diversification process of K-M526 likely occurred in Southeast Asia, with subsequent westward expansions of the ancestors of haplogroups R and Q."
Origins
The origins of haplogroup R1 remain unclear. It and its sibling clade R2 (R-M79) are the only immediate descendants of Haplogroup R (R-M207). R is a direct descendant of Haplogroup P1 (P-M45), and a sibling clade, therefore, of Haplogroup Q (Q-M242).
There were few areas in which Haplogroups P-M45, Q-M242 and R-M207 were all common amongst prehistoric populations. R-M207 and its subclades were most common along an axis from Western Europe to South Asia, whereas Q-M242 was the most common Y-DNA lineage among Native Americans. However, both P-M45 and its immediate descendants also appear to have been relatively common in Central Asia and Siberia.
Based on its ancestral lineages, an inferred origin for haplogroup R1 is South Asia or its western neighboring areas. For example, Kivisild 2003 believes the evidence "suggests that southern and western Asia might be the source of this haplogroup" and that "given the geographic spread and STR diversities of sister clades R1 and R2, the latter of which is restricted to India, Pakistan, Iran, and southern central Asia, it is possible that southern and western Asia were the source for R1 and R1a differentiation." Soares 2010 felt in their review of the literature, that the case for South Asian origins is strongest, with the Central Asian origin argued by (Wells 2001) being also worthy of consideration.
Karafet et al. (2014) "rapid diversification process of K-M526 likely occurred in Southeast Asia, with subsequent westward expansions of the ancestors of haplogroups R and Q."
Eurasia
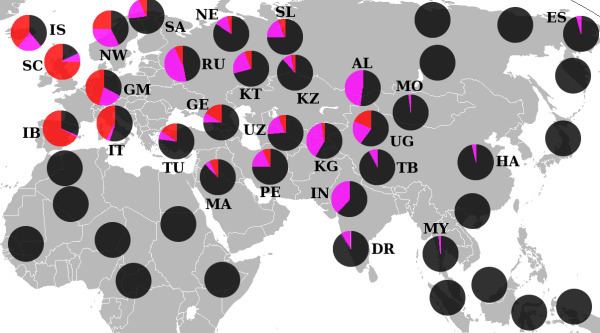
Haplogroup R1 is very common throughout all of Eurasia except East Asia and Southeast Asia. Its distribution is believed to be associated with the re-settlement of Eurasia following the last glacial maximum. Its main subgroups are R1a and R1b. One subclade of haplogroup R1b (especially R1b1a2), is the most common haplogroup in Western Europe and Bashkortostan (Lobov 2009), while a subclade of haplogroup R1a (especially haplogroup R1a1) is the most common haplogroup in large parts of South Asia, Eastern Europe, Central Asia, Western China, and South Siberia.
Individuals whose Y-chromosomes possess all the mutations on internal nodes of the Y-DNA tree down to and including M207 (which defines Haplogroup R) but which display neither the M173 mutation that defines haplogroup R1 nor the M479 mutation that defines Haplogroup R2 are categorized as belonging to group R* (R-M207). R* has been found in 10.3% (10/97) of a sample of Burusho and 6.8% (3/44) of a sample of Kalash from northern Pakistan (Firasat 2007).
Americas
The presence of haplogroup R1 among Indigenous Americans groups is a matter of controversy. It is now the most common haplogroup after the various Q-M242, especially in North America, highest worldwide R1 rates among Great Lakes/Algonquian-speakers, in Ojibwe people at 79%, Chipewyan 62%, Seminole 50%, Cherokee 47%, Dogrib 40% and Papago 38%. 97% of R1 had the M269 SNP (Single Nucleotide Polymorphism), which defines haplogroup R1b1b.
The poorly resolved SNP characterization led to much controversy. Some Native American Short Tandem Repeats haplotypes of R1, are not shared with Europeans. The evidence of decreasing gradient of haplogroup R from Northeastern to Southwestern North America further suggests that this results from European admixture.
Other authorities point to the greater similarity between haplogroup R1 subclades found in North America and those found in Siberia (e.g. Lell and Raghavan ), suggesting prehistoric immigration from Asia and/or Beringia, deriving from two major Siberian migrations. The first migration came from middle Siberia with the founding haplotype P-M45(x Q-M3). A second migration came from southeastern Siberia (Lower Amur/Sea of Okhkotsk/Kamchatka region) with the founding haplotype P-M45(x R1-M173), delineated by the RPS4Y-T marker, and took place at 7,000–9,500 years before present. Significant frequencies of RPS4Y-T are found in several northern Amerindian and Na-Dene populations (Bergen et al. 1999; Karafet et al. 1999), and in Lake Baikal region and Mongolia (Karafet et al. 1999), but is absent in Europeans (Bergen et al. 1999), reaching its highest frequencies in the populations of eastern Siberia. The P-M45(x R1-M173) subhaplogroup essentially seems to connect the population of eastern Siberians with the North- and Central-American Na-Dene and the surrounding Amerindian speakers. The RPS4Y-T diversification lends toward east Asia. The data correlates well with previous conclusions about the maternal migrations into Americas.
R1-positive P-M45 tested populations: Udegeys, Koryaks, North- and Central-American natives.
Africa
One subclade, now known as R1b1a2 (R-V88), is found only at high frequencies amongst populations native to West Africa, such as the Fulani, and is believed to reflect a prehistoric back-migration from Eurasia to Africa.
R1a (R-M420)
The highest levels of haplogroup R1a (>50%) are found amongst West Bengal Brahmins (72%), and Uttar Pradesh Brahmins, (67%), the Ishkashimi (68%), the Tajik population of Panjikent (64%), the Kyrgyz population of Central Kyrgyzstan (63.5%), Sorbs (63.39%), Bihar Brahmins (60.53%), Shors (58.8%), Poles (56.4%), Teleuts (55.3%), South Altaians (58.1%), Ukrainians (50%) and Russians (50%) (Semino 2000, Wells 2001, Behar 2003, and Sharma 2007).
R1a has been variously associated with:
The modern studies for R-M173 suggest that it could have originated in South Asia. It could have found its way initially from Western India (Gujarat) through Pakistan and Kashmir, then via Central Asia and Russia, before finally coming to Europe"..."as part of an archaeologically dated Paleolithic movement from east to west 30,000 years ago (Underhill 2009).
R1b (R-M343)
Haplogroup R1b probably originated in Eurasia prior to or during the last glaciation. It is the most common haplogroup in Western Europe and Bashkortostan.(Lobov 2009) It may have survived the last glacial maximum, in refugia near the southern Ural Mountains and Aegean Sea.(Lobov 2009).
It is also present at lower frequencies throughout Eastern Europe, with higher diversity than in western Europe, suggesting an ancient migration of haplogroup R1b from the east. Haplogroup R1b is also found at various frequencies in many different populations near the Ural Mountains and Central Asia, its likely region of origin.
There may be a correlation between this haplogroup and the spread of Centum branch Indo-European languages in southern and western Europe. For instance, the modern incidence of R1b reaches between 60% and 90% of the male population in most parts of Spain, Portugal, France, Britain and Ireland. It is also found in North Africa, where its frequency surpasses 10% in some parts of Algeria.
The R1b clade appears to have a much higher degree of internal diversity than R1a, which suggests that the M343 mutation that derives R1b from R-M173* may have occurred considerably earlier than the mutation that defines R1a.
Although it is rare in South Asia, some populations show relatively high percentages for R1b. These include Lambadi showing 37% (Kivisild 2005), Hazara 32% (Sengupta 2005), and Agharia (in East India) at 30% (Sengupta 2005). Besides these, R1b has appeared in Balochi (8%), Bengalis (6.5%), Chenchu (2%), Makrani (5%), Newars (10.6%), Pallan (3.5%) and Punjabis (7.6%) (Kivisild 2003, Sengupta 2005, and Gayden 2007).
R-M343 (previously called Hg1 and Eu18) is the most frequent Y-chromosome haplogroup in Europe. It is an offshoot of R-M173, characterised by the M343 marker. An overwhelming majority of members of R-M343 are classified as R-P25 (defined by the P25 marker), the remainder as R-M343*. Its frequency is highest in Western Europe (and due to modern European emigration, in parts of the Americas). The majority of R-M343-carriers of European descent belong to the R-M269 (R1b1a2) descendant line.
In popular culture
A fictional animation by Artem Lukichev links the history of R1, R1a and R1b to a traditional epic of the Bashkir people of the Ural Mountains.