
Ancestor IJ | Descendants I*, I1, I2 | |
 | ||
Possible time of origin Present 31-35,000 years BP Defining mutations L41, M170, M258, P19_1, P19_2, P19_3, P19_4, P19_5, P38, P212, U179 | ||
Haplogroup I (M170) is a Y-chromosome DNA haplogroup. It is a subgroup of haplogroup IJ, which itself is a derivative of the haplogroup IJK. I-M170 is one of the most numerous haplogroups among European males. It can be found in most present-day European populations, with peaks in some Northern European and South East European countries. Consequently, the haplogroup represents up to one-fifth of the male population of Europe, being the continent's second major Y-DNA haplogroup (behind Haplogroup R). Subclades of haplogroup I-M170 have also been found at high levels among some populations of the Caucasus.
Contents
- Origins
- Distribution
- Subgroups
- I M170
- I1 M253
- I2 M438
- I2a1a M26
- I2a1b M423
- I2a2 M436
- Specifications of mutation
- References
The haplogroup reaches its maximum frequency in the Balkans (with the highest concentration in present-day Herzegovina). It may be associated with unusually tall males, since those in the Dinaric Alps have been reported to be the tallest in the world, with an average height of 185.6 cm (6 ft 1.1 in).
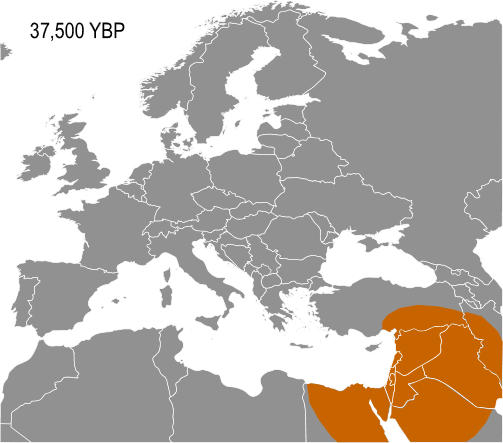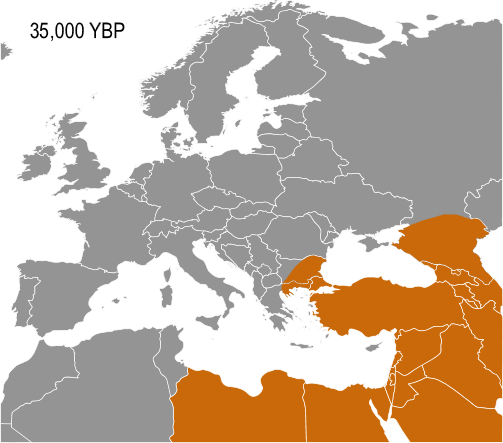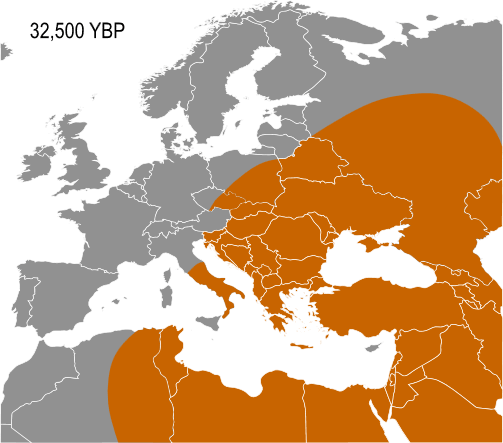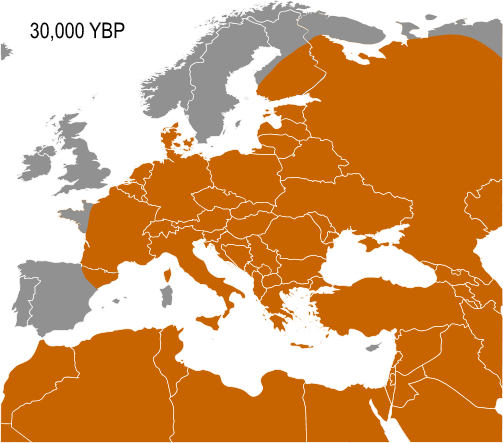
A European point of origin for I-M170 is generally proposed as it is not found outside of Europe on Paleolithic remains. However, the discovery in 2012 of living examples of Haplogroup IJ* in Mazanderani from Iran and Persians from Fars, may indicate that IJ originated in South West Asia.
Origins
Haplogroup IJ, carried by the Cro-Magnons was in the Middle East and/or Europe about 40,000 years ago. The TMRCA (time to most recent common ancestor) for I-M170 was estimated by Karafet and colleagues in 2008 to be 22,200 years ago, with a confidence interval between 15,300–30,000 years ago. This would make the founding event of I-M170 approximately contemporaneous with the Last Glacial Maximum (LGM), which lasted from 26,500 years ago until approximately 19,500 years ago. TMRCA is an estimate of the time of subclade divergence. Rootsi and colleagues in 2004 also note two other dates for a clade, age of STR variation, and time since population divergence. These last two dates are roughly associated, and occur somewhat after subclade divergence. For Haplogroup I-M170 they estimate time to STR variation as 24,000 ±7,100 years ago and time to population divergence as 23,000 ±7,700 years ago. These estimates are consistent with those of Karafet 2008 cited above. However Underhill and his colleagues calculate the time to subclade divergence of I1 and I2 to be 28,400 ±5,100 years ago, although they calculate the STR variation age of I1 at only 8,100 ±1,500 years ago.
Semino (2000) speculated that the initial dispersion of this population corresponds to the diffusion of the Gravettian culture. Later the haplogroup, along with two cases of Haplogroup C, was found on human remains belonging to the culture and on individuals of the Magdalenian and Azilian cultures. Rootsi and colleagues in 2004 suggested that each of the ancestral populations now dominated by a particular subclade of Haplogroup I-M170 experienced an independent population expansion immediately after the last glacial maximum.
The five known cases of Haplogroup I from Paleolithic European human remains make it the most frequent haplogroup from that period. In 2016, the 31,210–34,580 year old remains of a hunter-gatherer from Paglicci Cave, Apulia, Italy were found to carry I-M170. So far, only Haplogroup F* and Haplogroup C1b have been documented, once each, on older remains in Europe. I-M170 is the main haplogroup found on male remains in Mesolithic Europe, until circa 6,000 BCE.
Due to the arrival of so-called Early European Farmers, I-M170 is outnumbered by Haplogroup G among Neolithic European remains and by Haplogroup R in later remains.
The earliest documentation of I1 is from Neolithic Hungary, although it must have separated from I2 at an earlier point in time.
In one instance, haplogroup I was found far from Europe, among 2,000 year old remains from Mongolia.
As of 2015 researches, the earliest light eyes and light hair of hominid (Homo Sapiens) individuals after the long extinct Neandarthals have been documented at 8,000 years old remains in Motala, Sweden, belonging to subclades of Haplogroup I2 and mitochondrial Haplogroup U5. An I2a1 carrier was a carrier of red hair and others of genes of blond/light hair, while all the Motala hunter-gatherers were light skinned and blue eyed males. (Light skin genes, but not those for blonde/red hair have been found in: Siberia on a 17,000 year-old carrier of Haplogroup R*, as well as 8,000–9,000 year-old R1a remains from Karelia.)
It would seem to be that separate waves of population movement impacted Southeastern Europe. The role of the Balkans as a long-standing corridor to Europe from Anatolia and/or the Caucasus is shown by the common phylogenetic origins of both haplogroups I and J in the parent haplogroup IJ (M429). This common ancestry suggests that the subclades of IJ entered the Balkans from Anatolia or the Caucasus, some time before the Last Glacial Maximum. I and J were subsequently distributed in Asia and Europe in a disjunctive phylogeographic pattern typical of "sibling" haplogroups. A natural geographical corridor like the Balkans is likely to have been used later by members of other subclades of IJ, as well as other haplogroups, including those associated with Early European Farmers.
The existence of Haplogroup IJK – the ancestor of both haplogroups IJ and K (M9) – and its evolutionary distance from other subclades of Haplogroup F (M89), supports the inference that haplogroups IJ and K both arose in Southwestern Asia. Living carriers of F* and IJ* have been reported, from the Iranian plateau.
Distribution
Frequencies of Haplogroup I:
Subgroups
The subclades of Haplogroup I-M170 with their defining mutations:
Note that the naming of some of the subgroups has changed, as new markers have been identified, and the sequence of mutations has become clearer..
I-M170
The composite subclade I-M170 contains individuals directly descended from the earliest members of Haplogroup I, bearing none of the subsequent mutations which identify the remaining named subclades.
Several I* individuals, who do not fall into any known subclades, have been found among the Lak people of Dagestan, at a rate of (3/21), as well as Turkey (8/741), Adygea in the Caucasus (2/138) and Iraq (1/176), even though I-M170 occurs at only very low frequencies among modern populations of these regions as a whole. This is consistent with the belief that the haplogroup first appeared in South West Eurasia.
There are also high frequencies of Haplogroup I* among the Andalusians (3/103), French (4/179), Slovenians (2/55), Tabassarans (1/30), and Saami (1/35).
(Neither study from which the above figures were drawn excluded the present I2-M438 clade as a whole, but only certain subclades, so these presumed cases I* may possibly belong to I2.)
A living Hazara male from Afghanistan has also been found to carry I*, with all known subclades of both I1 (M253) and I2 (M438) ruled out.
I1-M253
Haplogroup I1-M253 (M253, M307, P30, P40) displays a very clear frequency gradient, with a peak frequency of approximately 35% among the populations of southern Norway, southwestern Sweden, and Denmark, and rapidly decreasing frequencies toward the edges of the historically Germanic-influenced world. A notable exception is Finland, where frequency in West Finns is up to 40%, and in certain provinces like Satakunta more than 50%.
Outside Fennoscandia, distribution of Haplogroup I1-M253 is closely correlated with that of Haplogroup I2a2-M436; but among Scandinavians (including both Germanic and Uralic peoples of the region) nearly all the Haplogroup I-M170 Y-chromosomes are I1-M253. Another characteristic of the Scandinavian I1-M253 Y-chromosomes is their rather low haplotype diversity (STR diversity): a greater variety of Haplogroup I1-M253 Y-chromosomes has been found among the French and Italians, despite the much lower overall frequency of Haplogroup I1-M253 among the modern French and Italian populations.
I2-M438
Haplogroup I2-M438, previously I1b, may have originated in southern Europe – it is now found at its highest frequencies in the western Balkans and Sardinia – some 15,000–17,000 years ago and developed into three main subgroups : I2-M438*, I2a-L460, I2b-L415 and I2c-L596.
I2a1a-M26
Haplogroup I2a1a-M26 is notable for its strong presence in Sardinia. Haplogroup I-M170 comprises approximately 40% of all patrilines among the Sardinians, and I2a1a-M26 is the predominant type of I among them.
Haplogroup I2a1a-M26 is practically absent east of France and Italy, while it is found at low but significant frequencies outside of Sardinia in the Balearic Islands, Castile-León, the Basque Country, the Pyrenees, southern and western France, and parts of the Maghreb in North Africa, Great Britain, and Ireland. Haplogroup I2a1a-M26 appears to be the only subclade of Haplogroup I-M170 found among the Basques, but appears to be found at somewhat higher frequencies among the general populations of Castile-León in Spain and Béarn in France than among the population of ethnic Basques. The M26 mutation is found in native males inhabiting every geographic region where megaliths may be found, including such far-flung and culturally disconnected regions as the Canary Islands, the Balearic Isles, Corsica, Ireland, and Sweden.
The distribution of I2a1a-M26 also mirrors that of the Atlantic Bronze Age cultures, which indicates a potential spread via the obsidian trade or a regular maritime exchange of some of metallurgical products.
I2a1b-M423
Haplogroup I2a1b-M423 is the most frequent Y-chromosome Haplogroup I-M170 in Central and Eastern European populations, reaching its peak in the Western Balkans, most notably in Dalmatia (50-60%) and Bosnia-Herzegovina (up to 71%, avg. 40-50%). A greater variance of this group has been found in Ireland and Great Britain, but overall frequency is very low (2-3%). Haplogroup I2a1b-M423 is virtually absent in Fennoscandia, Western and Southwestern Europe.
I2a2-M436
The distribution of Haplogroup I2a2-M436 (M436/P214/S33, P216/S30, P217/S23, P218/S32) is closely correlated to that of Haplogroup I1 except in Fennoscandia, which suggests that it was probably harbored by at least one of the Paleolithic refuge populations that also harbored Haplogroup I1-M253; the lack of correlation between the distributions of I1-M253 and I2a2-M436 in Fennoscandia may be a result of Haplogroup I2a2-M436's being more strongly affected in the earliest settlement of this region by founder effects and genetic drift due to its rarity, as Haplogroup I2a2-M436 comprises less than 10% of the total Y-chromosome diversity of all populations outside of Lower Saxony. Haplogroup I2a2-M436 has been found in over 4% of the population only in Germany, the Netherlands, Belgium, Denmark, England (not including Cornwall), Scotland, and the southern tips of Sweden and Norway in Northwest Europe; the provinces of Normandy, Maine, Anjou, and Perche in northwestern France; the province of Provence in southeastern France; the regions of Tuscany, Umbria, and Latium in Italy; and Moldavia and the area around Russia's Ryazan Oblast and Republic of Mordovia in Eastern Europe. One subclade of Haplogroup I2a2-M436, namely I2a2a1a1-M284, has been found almost exclusively among the population of Great Britain, which has been taken to suggest that the clade may have a very long history in that island. It is notable, however, that the distributions of Haplogroup I1-M253 and Haplogroup I2a2-M436 seem to correlate fairly well with the extent of historical influence of Germanic peoples. The punctual presence of both haplogroups at a low frequency in the area of the historical regions of Bithynia and Galatia in Turkey may be related to the Varangian Guard or rather suggests a connection with the ancient Gauls of Thrace, several tribes of which are recorded to have immigrated to those parts of Anatolia at the invitation of Nicomedes I of Bithynia. This suggestion is supported by recent genetic studies regarding Y-DNA Haplogroup I2b2-L38 have concluded that there was some Late Iron Age migration of Celtic La Tène people, through Belgium, to the British Isles including north-east Ireland.
Haplogroup I2a2-M436 also occurs among approximately 1% of Sardinians, and in Hazaras from Afghanistan at 3%.
Specifications of mutation
The technical details of U179 are:
Nucleotide change (rs2319818): G to APosition (base pair): 275Total size (base pairs): 220Forward 5′→ 3′: aaggggatatgacgactgattReverse 5′→ 3′: cagctcctcttttcaactctca