
Possible time of origin approx 24,100 years BP Ancestor E-M215/M35 Defining mutations Z827 | Possible place of origin Horn of Africa Descendants E-L19, E-Z830 | |
 | ||
E-Z827, also known as E1b1b1b, is a major human Y chromosome haplogroup. It is the parent lineage to the E-Z830 and E-V257 subclades, and defines their common ancestry. The former is predominantly found in the Horn of Africa and the Middle East, whereas the latter is most frequently observed in Northwestern Africa. It is also found at lower frequencies in Europe, and in isolated parts of Southeast Africa.
Contents
Family Tree
The following phylogeny is based on the YCC 2008 tree and subsequent published research as summarized by ISOGG.
E-V257/L19 (E1b1b1b1)
E-V257/L19 showed a parallel with its sibling clade E-V68 in the way that both clades show signs of having migrated from North Africa to southern Europe across the Mediterranean sea. 6 "E-V257/L19*" individuals were found in published samples who were E-V257/L19, but not E-M81. a Marrakesh Berber, a Corsican, a Sardinian, A Borana from Kenya, a southern Spaniard and a Cantabrian.
Within E-M35, there are striking parallels between two haplogroups, E-V68 and E-V257. Both contain a lineage which has been frequently observed in North Africa (E-M78 and E-M81, respectively) and a group of undifferentiated chromosomes that are mostly found in southern Europe. An expansion of E-M35 carriers, possibly from the Middle East as proposed by other authors, and split into two branches separated by the geographic barrier of the Mediterranean Sea, would explain this geographic pattern. However, the absence of E-V68* and E-V257* in the Middle East makes a maritime spread between northern Africa and southern Europe a more plausible hypothesis.
A project dedicated to researching and understanding the origins of E-V257/L19* is underway at FamilyTreeDNA.com. The name of the project is E1b1b1b*-A
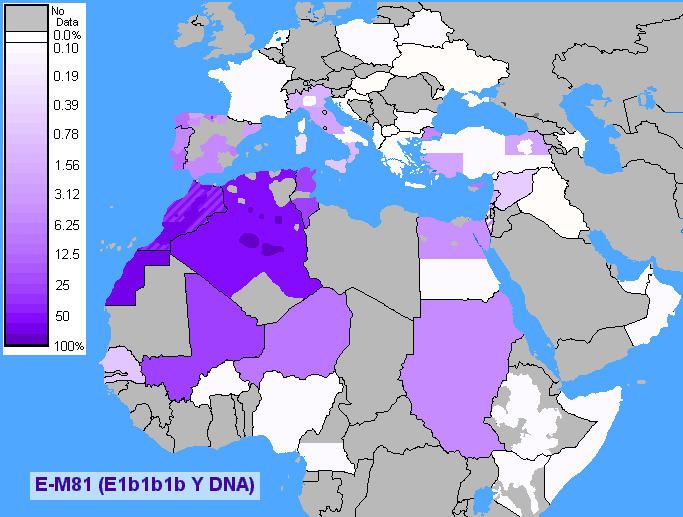
E-V257's dominant sub-clade E-M81 is thought to have originated in the area of North Africa 14,200 years ago.
E-M81
E-M81 is the most common subclade of E-L19/V257 and found in the Maghreb, dominated by its sub-clade E-M183. This haplogroup reaches a mean frequency of 42% in North Africa, decreasing in frequency from 100% in some isolated Berber populations to approximately 10% to the east of this range in Egypt. Because of its prevalence among these groups and also others such as Mozabite, Middle Atlas, Kabyle and other Berber groups, it is sometimes referred to as a genetic "Berber marker". Pereira et al. (2010) report high levels among Tuareg in two Saharan populations - 77.8% near Gorom-Gorom, in Burkina Faso, and 81.8% from Gossi in Mali. There was a much lower frequency of 11.1% in the vicinity of Tanut in the Republic of Niger.
E-M81 is also quite common among North African Arabic-speaking groups. It is generally found at frequencies around 45% in coastal cities of the Maghreb (Oran, Tunis, Tizi Ouzou, Algiers).
In this key area from Egypt to the Atlantic Ocean, Arredi et al. (2004) report a pattern of decreasing STR haplotype variation (implying decreasing lineage age in those areas) from East to West, accompanied by a substantial increasing frequency. At the eastern extreme of this core range, Kujanova et al. (2009) found M81 in 28.6% (10 out of 35 men) in El-Hayez in the Western desert in Egypt
Arredi et al. (2004) believe the pattern of distribution and variance to be consistent with the hypothesis of a post Paleolithic "demic diffusion" from the East. The ancestral lineage of E-M81 in their hypothesis could have been linked with the spread of Neolithic food-producing technologies from the Fertile Crescent via the Nile, although pastoralism rather than agriculture. E-M81 and possibly proto-Afroasiatic language may have been carried either all the way from Asia, or they may represent a "local contribution to the North African Neolithic transition". According to Shomarka Keita, a Near Eastern origin of proto-Afroasiatic speakers carrying E-M81, or its ancestral lineage, is inconsistent with the linguistic evidence, which seems to indicate an African origin of Proto-Afro-Asiatic speakers. Keita argues that there is no autochthonous presence of E-M81 in the Near East, indicating that M81 most likely emerged from its parent clade M35 either in the Maghreb, or possibly as far southeast as the Horn of Africa.
The E-M81 subclade has been found in ancient Guanche (Bimbapes) fossils excavated in Punta Azul, El Hierro, Canary Islands, which are dated to the 10th century (~44%).
Europe
In Europe, E-M81 has a widespread distribution at very low frequencies but is common mostly in the Iberian Peninsula, where unlike in the rest of Europe it is more common than E-M78, with an average frequency around 5%. Its frequencies are higher in the western half of the peninsula with frequencies reaching 8% in Extremadura and South Portugal, 4% in one study and 9% in another in Galicia, 10% in Western Andalusia and Northwest Castile and 9% to 17% in Cantabria. The highest frequencies of this clade found so far in Europe were observed in the Pasiegos from Cantabria, ranging from 18% (8/45) to 41% (23/56). An average frequency of 8.28% (54/652) has also been reported in the Spanish Canary Islands with frequencies over 10% in the three largest islands of Tenerife (10.68%), Gran Canaria (11.54%) and Fuerteventura (13.33%).
E-M81 is also found in France, 2.70% (15/555) overall with frequencies surpassing 5% in Auvergne (5/89) and Île-de-France (5/91), in Sicily (approximately 2% overall, but up to 7% in Piazza Armerina), and in slightly lower frequencies in continental Italy (especially near Lucera) due to historic colonization during the Islamic, Roman, and Carthaginian empires or ancient migrations in the Metals Ages through maritime means. E-M81 was also found in 2013 at 5.8% in a large sample of 1 204 Sardinians.
Latin America
As a result of Spanish and Portuguese colonization of the Americas, this sub-clade is found throughout Latin America, for example 6.1% in Cuba, 5.4% in Brazil (Rio de Janeiro), and among Hispanic men from California and Hawaii 2.4%.
Others
In smaller numbers, E-M81 men can be found in areas in contact with the Maghreb, both around the Sahara, in places like Sudan, and around the Mediterranean in places like Lebanon, Turkey, and amongst Sephardic Jews.
Distribution
The following gives a summary of most of the studies which specifically tested for E-M81, showing where its distribution is greater than 1% in Europe, North Africa, the Middle East and Latin America.
E-Z830 (E1b1b1b2)
A recently confirmed sub-clade of E-Z827, Z830, includes the confirmed sub-clades of E-M123, E-M293, and E-V42, and is a sibling clade to E-L19. Currently, the E-M35 phylogeny project recognizes four distinct clusters of Z830* carriers, two of which are exclusively Jewish in origin. The remaining two are significantly smaller, and include scattered individuals in Germany, Spain, Latin America, Egypt, and Ethiopia.
E-M123
E-M123 is mostly known for its major subclade E-M34, which dominates this clade.
E-V1515
A new clade (E-V1515) was defined by Trombetta et al. 2015, which originated about 12 kya (95% CI 8.6-16.4) in eastern Africa where it is currently mainly distributed. This clade includes all the sub-Saharan haplogroups (E-V42, E-M293, E-V92, E-V6) reported as E-M35 basal clades in a previous phylogeny.
E-M293
E-M293 is a subclade of E-V1515. It was identified by ISOGG as the second clade within E-Z830. It was discovered before E-Z830, being announced in Henn 2008, which associated it with the spread of pastoralism from Eastern Africa into Southern Africa. So far high levels have been found in specific ethnic groups in Tanzania and Southern Africa. Highest were the Datog (43%), Khwe (Kxoe) (31%), Burunge (28%), and Sandawe (24%). Henn (2008) in their study also found two Bantu-speaking Kenyan males with the M293 mutation. Other E-M215 subclades are rare in Southern Africa. The authors state...
Without information about M293 in the Maasai, Hema, and other populations in Kenya, Sudan, and Ethiopia, we cannot pinpoint the precise geographic source of M293 with greater confidence. However, the available evidence points to present-day Tanzania as an early and important geographic locus of M293 evolution.
They also say that "M293 is only found in sub-Saharan Africa, indicating a separate phylogenetic history for M35.1 * (former) samples further north". E-P72 appears in Karafet (2008). Trombetta et al. 2011 announced that this is a subclade of E-M293.
E-V42
Trombetta et al. 2011 announced the discovery of E-V42 in two Ethiopian Jews. It was suggested that it may be restricted to the region around Ethiopia. However, further testing by commercial DNA testing companies confirmed positive results for this subclade in Arabia as well.
E-V6
The E-V6 subclade of E-V1515 is defined by V6. Cruciani et al. (2004) identified a significant presence of these lineages in Ethiopia, and also some in the neighboring Somali population. Among the Ethiopian and Somali samples, the highest were 14.7% among the Ethiopian Amhara, and 16.7% among the Ethiopian Wolayta.
E-V92
Trombetta et al. 2011 announced the discovery of E-V92 in two Ethiopian Amhara. Like E-V6 and E-V42 it possibly only exists in the area of Ethiopia.
Phylogenetic History
Prior to 2002, there were in academic literature at least seven naming systems for the Y-Chromosome Phylogenetic tree. This led to considerable confusion. In 2002, the major research groups came together and formed the Y-Chromosome Consortium (YCC). They published a joint paper that created a single new tree that all agreed to use. Later, a group of citizen scientists with an interest in population genetics and genetic genealogy formed a working group to create an amateur tree aiming at being above all timely. The table below brings together all of these works at the point of the landmark 2002 YCC Tree. This allows a researcher reviewing older published literature to quickly move between nomenclatures.
Original Research Publications
The following research teams per their publications were represented in the creation of the YCC Tree.