
Symbol Kinesin motor domain InterPro IPR001752 PROSITE PS50067 | Pfam PF00225 SMART SM00129 SCOP 1bg2 | |
 | ||
A kinesin is a protein belonging to a class of motor proteins found in eukaryotic cells.
Contents
- The kinesins
- Overall structure
- Kinesin motor domain
- Cargo transport
- Direction of motion
- Proposed mechanisms of movement
- Theoretical modeling of kinesin
- Kinesin and mitosis
- Kinesin superfamily members
- References
Kinesins move along microtubule (MT) filaments, and are powered by the hydrolysis of adenosine triphosphate (ATP) (thus kinesins are ATPases). The active movement of kinesins supports several cellular functions including mitosis, meiosis and transport of cellular cargo, such as in axonal transport. Most kinesins walk towards the positive end of a microtubule, which, in most cells, entails transporting cargo such as protein and membrane components from the centre of the cell towards the periphery. This form of transport is known as anterograde transport. In contrast, dyneins are motor proteins that move toward the microtubules' negative end.
The kinesins
Kinesins were discovered as MT-based anterograde intracellular transport motors. The founding member of this superfamily, kinesin-1, was isolated as a heterotetrameric fast axonal organelle transport motor consisting of 2 identical motor subunits (KHC) and 2 "light chains" (KLC) via microtubule affinity purification from neuronal cell extracts. Subsequently, a different, heterotrimeric plus-end-directed MT-based motor named kinesin-2, consisting of 2 distinct KHC-related motor subunits and an accessory "KAP" subunit, was purified from echinoderm egg/embryo extracts and is best known for its role in transporting protein complexes (IFT particles) along axonemes during cilium biogenesis. Molecular genetic and genomic approaches have led to the recognition that the kinesins form a diverse superfamily of motors that are responsible for multiple intracellular motility events in eukaryotic cells. For example, the genomes of mammals encode more than 40 kinesin proteins, organized into at least 14 families named kinesin-1 through kinesin-14.
Overall structure
Members of the kinesin superfamily vary in shape but the prototypical kinesin-1 is a heterotetramer whose motor subunits (heavy chains or KHCs) form a protein dimer (molecule pair) that binds two light chains (KLCs).
The heavy chain of kinesin-1 comprises a globular head (the motor domain) at the amino terminal end connected via a short, flexible neck linker to the stalk – a long, central alpha-helical coiled coil domain – that ends in a carboxy terminal tail domain which associates with the light-chains. The stalks of two KHCs intertwine to form a coiled coil that directs dimerization of the two KHCs. In most cases transported cargo binds to the kinesin light chains, at the TPR motif sequence of the KLC, but in some cases cargo binds to the C-terminal domains of the heavy chains.
Kinesin motor domain
The head is the signature of kinesin and its amino acid sequence is well conserved among various kinesins. Each head has two separate binding sites: one for the microtubule and the other for ATP. ATP binding and hydrolysis as well as ADP release change the conformation of the microtubule-binding domains and the orientation of the neck linker with respect to the head; this results in the motion of the kinesin. Several structural elements in the Head, including a central beta-sheet domain and the Switch I and II domains, have been implicated as mediating the interactions between the two binding sites and the neck domain. Kinesins are structurally related to G proteins, which hydrolyze GTP instead of ATP. Several structural elements are shared between the two families, notably the Switch I and Switch II domains.
Cargo transport
In the cell, small molecules such as gases and glucose diffuse to where they are needed. Large molecules synthesised in the cell body, intracellular components such as vesicles and organelles such as mitochondria (the powerhouse of the cell) are too large (and the cytosol too crowded) to be able to diffuse to their destinations. Motor proteins fulfill the role of transporting large cargo about the cell to their required destinations. Kinesins are motor proteins that transport such cargo by walking unidirectionally along microtubule tracks hydrolysing one molecule of adenosine triphosphate (ATP) at each step. It was thought that ATP hydrolysis powered each step, the energy released propelling the head forwards to the next binding site. However, it has been proposed that the head diffuses forward and the force of binding to the microtubule is what pulls the cargo along. In addition viruses, HIV for example, exploit kinesins to allow virus particle shuttling after assembly.
There is significant evidence that cargoes in-vivo are transported by multiple motors.
Direction of motion
Motor proteins travel in a specific direction along a microtubule. This is because the microtubule is polar and the heads only bind to the microtubule in one orientation, while ATP binding gives each step its direction through a process known as neck linker zippering.
Most kinesins walk towards the plus-end of a microtubule which, in most cells, entails transporting cargo from the center of the cell towards the periphery. This form of transport is known as anterograde transport/orthrograde transport. Kinesin-14 family proteins, such as Drosophila melanogaster NCD, budding yeast KAR3, and Arabidopsis thaliana ATK5, walk in the opposite direction, toward microtubule minus ends.
A different type of motor protein known as dyneins, move towards the minus end of the microtubule. Thus they transport cargo from the periphery of the cell towards the centre, for example from the terminal boutons of a neuronal axon to the cell body (soma). This is known as retrograde transport.
Cin8, a member of the Kinesin-5 family, has the novel ability to switch directionality. It has been shown to be minus-end-directed (contrary to the rest of the known Kinesins) when bound to a single microtubule, but plus-end-directed when cross-linking antiparallel microtubules (pushing the minus ends further apart and pulling the plus ends towards each other). This dual directionality has been observed in identical conditions where free Cin8 molecules move towards the minus end, but cross-linking Cin8 move toward the plus ends of each cross-linked microtubule. It is suggested that this unique ability is a result of coupling with other Cin8 motors and helps to fulfill the role of dynein in budding yeast.
Proposed mechanisms of movement
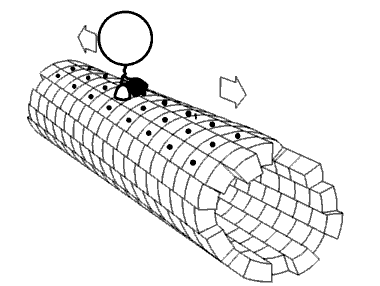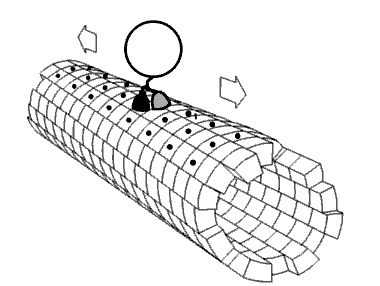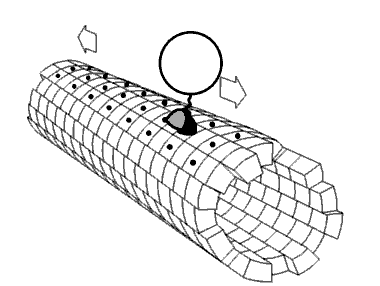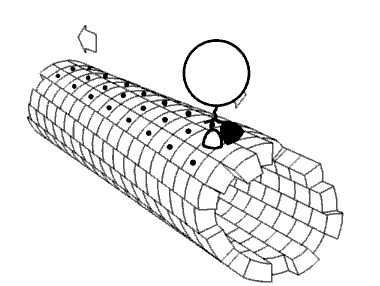
Kinesin accomplishes transport by "walking" along a microtubule. Two mechanisms have been proposed to account for this movement.
Despite some remaining controversy, mounting experimental evidence points towards the hand-over-hand mechanism as being more likely.
ATP binding and hydrolysis cause kinesin to travel via a "seesaw mechanism" about a pivot point. This seesaw mechanism accounts for observations that the binding of the ATP to the no-nucleotide, microtubule-bound state results in a tilting of the kinesin motor domain relative to the microtubule. Critically, prior to this tilting the neck linker is unable to adopt its motor-head docked, forward-facing conformation. The ATP-induced tilting provides the opportunity for the neck linker to dock in this forward-facing conformation. This model is based on CRYO-EM models of the microtubule-bound kinesin structure which represent the beginning and end states of the process, but cannot resolve the precise details of the transition between the structures.
Theoretical modeling of kinesin
A number of theoretical models of the molecular motor protein kinesin have been proposed. Many challenges are encountered in theoretical investigations given the remaining uncertainties about the roles of protein structures, precise way energy from ATP is transformed into mechanical work, and the roles played by thermal fluctuations. This is a rather active area of research. There is a need especially for approaches which better make a link with the molecular architecture of the protein and data obtained from experimental investigations.
The single-molecule dynamics are already well described but it seems that these nano scale machines typically work in large teams. Recent experimental research has shown that kinesins, while moving along microtubules, interact with each other, the interactions being short range and weak attractive (1.6±0.5 KBT). One model that has been developed takes into account these particle interactions, where the dynamic rates change accordingly with the energy of interaction. If the energy is positive the rate of creating bonds (q) will be higher while the rate of breaking bonds (r) will be lower. One can understand that the rates of entrance and exit in the microtubule will be changed as well by the energy (See figure 1 in reference 30). If the second site is occupied the rate of entrance will be α*q and if the last but one site is occupied the rate of exit will be β*r. This theoretical approach agrees with the results of Monte Carlo simulations for this model, especially for the limiting case of very large negative energy. The normal totally asymmetric simple exclusion process for (or TASEP) results can be recovered from this model making the energy equal to zero.
Kinesin and mitosis
In recent years, it has been found that microtubule-based molecular motors (including a number of kinesins) have a role in mitosis (cell division). Kinesins are important for proper spindle length and are involved in sliding microtubules apart within the spindle during prometaphase and metaphase, as well as depolymerizing microtubule minus ends at centrosomes during anaphase. Specifically, Kinesin-5 family proteins act within the spindle to slide microtubules apart, while the Kinesin 13 family act to depolymerize microtubules.
Kinesin superfamily members
Human kinesin superfamily members include the following proteins, which in the standardized nomenclature developed by the community of kinesin researchers, are organized into 14 families named kinesin-1 through kinesin-14:
kinesin-1 light chains:
kinesin-2 associated protein: