
Higher classification Chromalveolata | Scientific name Alveolata Rank Superphylum | |
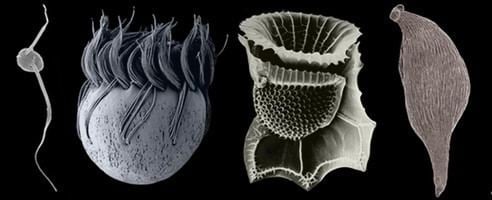 | ||
Lower classifications Dinoflagellate, Ciliate, Apicomplexa, Coccidia | ||
Alveolates
The alveolates (meaning "with cavities") are a group of protists, considered a major clade and superphylum within Eukarya, and are also called Alveolata.
Contents
- Alveolates
- Alveolate protozoa
- Characteristics
- Classification
- Phylogeny
- Taxonomy
- Development
- Evolution
- References
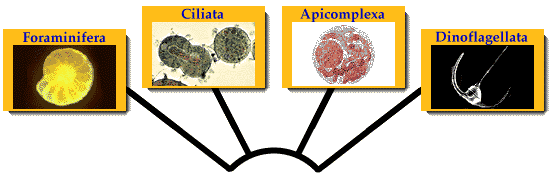
Alveolate protozoa
Characteristics
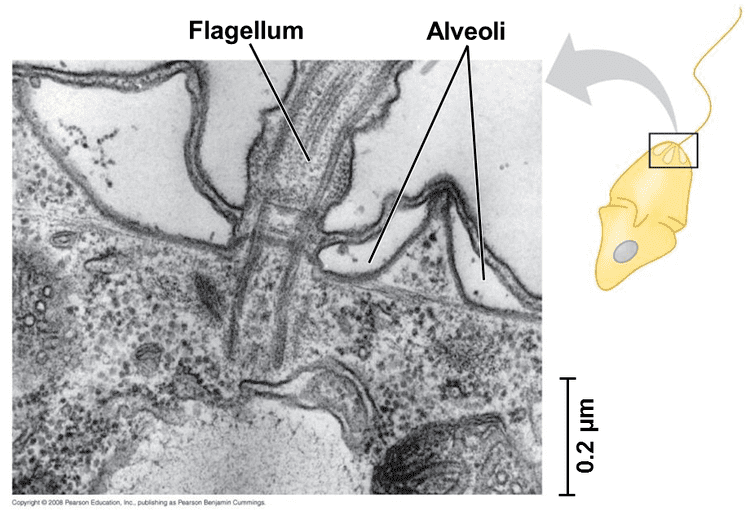
The most notable shared characteristic is the presence of cortical alveoli, flattened vesicles packed into a continuous layer supporting the membrane, typically forming a flexible pellicle. In Dinoflagellates they often form armor plates. Alveolates have mitochondria with tubular cristae and their flagella or cilia have a distinct structure.
Almost all sequenced mitochondrial genomes of Ciliates and apicomplexia are linear. The mitochondrial genome of Babesia microti is circular. This species is also now known not to belong to either of the genera Babesia or Theileria and a new genus will have to be created for it.
Classification
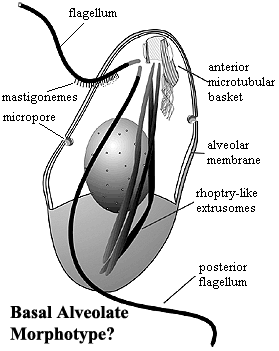
Alveolata comprises around 9 major and minor groups, which are very diverse in form, and are known to be related by various ultrastructural and genetic similarities:
The Acavomonidia and Colponemidia were previously grouped together as colponemids, a taxon now split based on ultrastructural analysis. The Acavomonidia are closer to the dinoflagellate/perkinsid group than the Colponemidia are. As such, the informal term "colponemids", as its stands currently, covers two non-sister groups within Alveolata: the Acavomonidia and the Colponemidia.
The Apicomplexa and dinoflagellates may be more closely related to each other than to the ciliates. Both have plastids, and most share a bundle or cone of microtubules at the top of the cell. In apicomplexans this forms part of a complex used to enter host cells, while in some colorless dinoflagellates it forms a peduncle used to ingest prey. Various other genera are closely related to these two groups, mostly flagellates with a similar apical structure. These include free-living members in Oxyrrhis and Colponema, and parasites in Perkinsus, Parvilucifera, Rastrimonas and the ellobiopsids. In 2001, direct amplification of the rRNA gene in marine picoplankton samples revealed the presence of two novel alveolate linages, called group I and II. Group I has no cultivated relatives, while group II is related to the dinoflagellate parasite Amoebophrya, which was classified until now in the Syndiniales dinoflagellate order.
Relationships between some of these the major groups were suggested during the 1980s, and a specific relationship between all three was confirmed in the early 1990s by genetic studies, most notably by Gajadhar et al. Cavalier-Smith, introduced the formal name Alveolata in 1991, although at the time he actually considered the grouping to be a paraphyletic assemblage, rather than a monophyletic group.
Some studies suggested the haplosporids, mostly parasites of marine invertebrates, might belong here but they lack alveoli and are now placed among the Cercozoa.
Phylogeny
Based on a compilation of the following works.
Taxonomy
Alveolata Cavalier-Smith 1991 [Alveolatobiontes]
Development
The development of plastids among the alveolates is intriguing. Cavalier-Smith proposed the alveolates developed from a chloroplast-containing ancestor, which also gave rise to the Chromista (the chromalveolate hypothesis). Other researchers have speculated that the alveolates originally lacked plastids and possibly the dinoflagellates and Apicomplexa acquired them separately. However, it now appears that the alveolates, the dinoflagellates, the Chromerida and the heterokont algae acquired their plastids from a red algae with evidence of a common origin of this organelle in all these four clades.
Evolution
A Bayesian estimate places the evolution of the alveolate group at ~850 million years ago. The Alveolata consist of Myzozoa, Ciliates, and Colponemids. In other words, The term Myzozoa meaning "to siphon the contents from prey", may be applied informally to the common ancestor of the subset of alveolates that are neither ciliates nor colponemids. Predation upon algae is an important driver in alveolate evolution, as it can provide sources for endosymbiosis of novel plastids. The term Myzozoa is therefore a handy concept for tracking the history of the alveolate phylum.
The ancestors of the alveolate group may have been photosynthetic. The ancestral alveolate probably possessed a plastid. Chromerids, apicomplexans, and peridinin dinoflagellates have retained this organelle. Going one step even further back, the chromerids, the peridinin dinoflagellates and the heterokont algae possess a monophyletic plastid lineage in common, i.e. acquired their plastids from a red alga, and so it seems likely that the common ancestor of alveolates and heterokonts was also photosynthetic.
In one school of thought the common ancestor of the dinoflagellates, apicomplexans, Colpodella, Chromerida, and Voromonas was a myzocytotic predator with two heterodynamic flagella, micropores, trichocysts, rhoptries, micronemes, a polar ring and a coiled open sided conoid. While the common ancestor of alveolates may also have possessed some of these characteristics, it has been argued that Myzocytosis was not one of these characteristics, as ciliates ingest prey by a different mechanism.
An ongoing debate concerns the number of membranes surrounding the plastid across apicomplexans and certain dinoflagellates, and the origin of these membranes. This ultrastructural character can be used to group organisms and if the character is in common, it can imply that phyla had a common photosynthetic ancestor. On the basis that apicomplexans possess a plastid surrounded by 4 membranes, and that peridinin dinoflagellates possess a plastid surrounded by 3 membranes, Petersen et al. have been unable to rule our that the shared stramenopile-alveolate plastid could have been recycled multiple times in the alveolate phylum, the source being stramenopile-alveolate donors, through the mechanism of ingestion and endosymbiosis.
Ciliates are a model apicomplexan, having been genetically studied in great depth over the longest period of any apicomplexan lineage. They are unusual among eukaryotes in that reproduction involves a micronucleus and a macronucleus. Their reproduction is easily studied in the lab, and made them a model eukaryote historically. Being entirely predatory and lacking any remnant plastid, their development as a phylum illustrates how predation and autotrophy are in dynamic balance and that the balance can swing one way or other at the point of origin of a new phylum from mixotrophic ancestors, causing one ability to be lost.