
TH H1.00.01.1.01032 | FMA 67472 | |
 | ||
A flagellum (/fləˈdʒɛləm/; plural: flagella) is a lash-like appendage that protrudes from the cell body of certain prokaryotic and eukaryotic cells. The word flagellum in Latin means whip. The primary role of the flagellum is locomotion, but it also often has function as a sensory organelle, being sensitive to chemicals and temperatures outside the cell. Flagella are organelles defined by function rather than structure. Large differences occur between different types of flagella; the prokaryotic and eukaryotic flagella differ greatly in protein composition, structure, and mechanism of propulsion. However, both can be used for swimming.
Contents
- Types
- Structure and composition
- Motor
- Assembly
- Evolution
- Flagella and the intelligent design debate
- Flagellar arrangement schemes
- Archaeal
- Terminology
- Internal structure
- Mechanism
- Flagella vs cilia
- Intraflagellar transport
- Evolution and occurrence
- Typology
- References
An example of a flagellated bacterium is the ulcer-causing Helicobacter pylori, which uses multiple flagella to propel itself through the mucus lining to reach the stomach epithelium. An example of a eukaryotic flagellate cell is the mammalian sperm cell, which uses its flagellum to propel itself through the female reproductive tract. Eukaryotic flagella are structurally identical to eukaryotic cilia, although distinctions are sometimes made according to function and/or length. Fimbriae and pili are also thin appendages, but have different functions and are usually smaller.
Types
Three types of flagella have so far been distinguished: bacterial, archaeal, and eukaryotic.
The main differences among these three types are:
Structure and composition
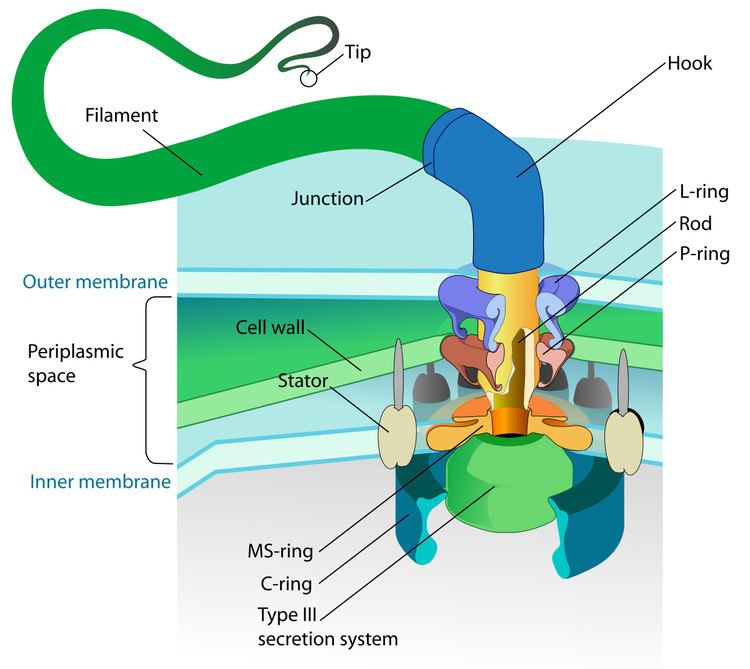
The bacterial flagellum is made up of the protein flagellin. Its shape is a 20-nanometer-thick hollow tube. It is helical and has a sharp bend just outside the outer membrane; this "hook" allows the axis of the helix to point directly away from the cell. A shaft runs between the hook and the basal body, passing through protein rings in the cell's membrane that act as bearings. Gram-positive organisms have two of these basal body rings, one in the peptidoglycan layer and one in the plasma membrane. Gram-negative organisms have four such rings: the L ring associates with the lipopolysaccharides, the P ring associates with peptidoglycan layer, the M ring is embedded in the plasma membrane, and the S ring is directly attached to the plasma membrane. The filament ends with a capping protein.
The flagellar filament is the long, helical screw that propels the bacterium when rotated by the motor, through the hook. In most bacteria that have been studied, including the Gram-negative Escherichia coli, Salmonella typhimurium, Caulobacter crescentus, and Vibrio alginolyticus, the filament is made up of 11 protofilaments approximately parallel to the filament axis. Each protofilament is a series of tandem protein chains. However, Campylobacter jejuni has seven protofilaments.
The basal body has several traits in common with some types of secretory pores, such as the hollow, rod-like "plug" in their centers extending out through the plasma membrane. Given the structural similarities between bacterial flagella and bacterial secretory systems, bacterial flagella may have evolved from the type-three secretion system; however, whether these pores are derived from the bacterial flagella or the bacterial secretory system is not known for certain.
Motor
The bacterial flagellum is driven by a rotary engine (Mot complex) made up of protein, located at the flagellum's anchor point on the inner cell membrane. The engine is powered by proton motive force, i.e., by the flow of protons (hydrogen ions) across the bacterial cell membrane due to a concentration gradient set up by the cell's metabolism (Vibrio species have two kinds of flagella, lateral and polar, and some are driven by a sodium ion pump rather than a proton pump). The rotor transports protons across the membrane, and is turned in the process. The rotor alone can operate at 6,000 to 17,000 rpm, but with the flagellar filament attached usually only reaches 200 to 1000 rpm. The direction of rotation can be switched almost instantaneously, caused by a slight change in the position of a protein, FliG, in the rotor. The flagellum is highly energy efficient and uses very little energy.
The cylindrical shape of flagella is suited to locomotion of microscopic organisms; these organisms operate at a low Reynolds number, where the viscosity of the surrounding water is much more important than its mass or inertia.
The rotational speed of flagella varies in response to the intensity of the proton motive force, thereby permitting certain forms of speed control, and also permitting some types of bacteria to attain remarkable speeds in proportion to their size; some achieve roughly 60 cell lengths per second. At such a speed, a bacterium would take about 245 days to cover 1 km; although that may seem slow, the perspective changes when the concept of scale is introduced. In comparison to macroscopic life forms, it is very fast indeed when expressed in terms of number of body lengths per second. A cheetah, for example, only achieves about 25 body lengths per second.
Through use of their flagella, E. coli is able to move rapidly towards attractants and away from repellents, by means of a biased random walk, with 'runs' and 'tumbles' brought about by rotating its flagellum counterclockwise and clockwise, respectively.
Assembly
During flagellar assembly, components of the flagellum pass through the hollow cores of the basal body and the nascent filament. During assembly, protein components are added at the flagellar tip rather than at the base. In vitro, flagellar filaments assemble spontaneously in a solution containing purified flagellin as the sole protein.
Evolution
At least 10 protein components of the bacterial flagellum share homologous proteins with the type-three secretion system (TTSS), hence one likely evolved from the other. Because the TTSS has a similar number of components as a flagellar apparatus (about 25 proteins), which one evolved first is difficult to determine. However, the flagellar system appears to involve more proteins overall, including various regulators and chaperones, hence it has been argued that flagella evolved from a TTSS. However, it has also been suggested that the flagellum may have evolved first or the two structures evolved in parallel. Early single-cell organisms' need for motility (mobility) support that the more mobile flagella would be selected by evolution first, but the TTSS evolving from the flagellum can be seen as 'reductive evolution', and receives no topological support from the phylogenetic trees. The hypothesis that the two structures evolved separately from a common ancestor accounts for the protein similarities between the two structures, as well as their functional diversity.
Flagella and the intelligent design debate
Some authors have argued that flagella cannot have evolved because they can only function properly when all proteins are in place. In other words, the flagellar apparatus is "irreducibly complex". This has long been debunked, because many proteins can be deleted or mutated and the flagellum still works, though sometimes at reduced efficiency. In fact, the composition of flagella is surprisingly diverse across bacteria, with many proteins only found in some species, but not others. Hence, the flagellar apparatus is clearly very flexible in evolutionary terms and perfectly able to lose or gain protein components.
Flagellar arrangement schemes
Different species of bacteria have different numbers and arrangements of flagella.
In certain large forms of Selenomonas, more than 30 individual flagella are organized outside the cell body, helically twining about each other to form a thick structure (easily visible with the light microscope) called a "fascicle".
Other bacteria, such as most spirochetes, have two or more specialized flagella (endoflagella) arising from opposite poles of the cell, which together constitute the so-called "axial filament" that is located within the periplasmic space between the flexible cell wall and an outer sheath. The rotation of the axial filament relative to the cell body causes the entire bacterium to move forward in a corkscrew-like motion, even through material viscous enough to prevent the passage of normally flagellated bacteria.
Counterclockwise rotation of a monotrichous polar flagellum pushes the cell forward with the flagellum trailing behind, much like a corkscrew moving inside cork. Indeed, water on the microscopic scale is highly viscous, very different from our daily experience of water.
Flagella are left-handed helices, and bundle and rotate together only when rotating counterclockwise. When some of the rotors reverse direction, the flagella unwind and the cell starts "tumbling". Even if all flagella would rotate clockwise, they likely will not form a bundle, due to geometrical, as well as hydrodynamic reasons. Such "tumbling" may happen occasionally, leading to the cell seemingly thrashing about in place, resulting in the reorientation of the cell. The clockwise rotation of a flagellum is suppressed by chemical compounds favorable to the cell (e.g. food), but the motor is highly adaptive to this. Therefore, when moving in a favorable direction, the concentration of the chemical attractant increases and "tumbles" are continually suppressed; however, when the cell's direction of motion is unfavorable (e.g., away from a chemical attractant), tumbles are no longer suppressed and occur much more often, with the chance that the cell will be thus reoriented in the correct direction.
In some Vibrio spp. (particularly Vibrio parahemolyticus) and related proteobacteria such as Aeromonas, two flagellar systems co-exist, using different sets of genes and different ion gradients for energy. The polar flagella are constitutively expressed and provide motility in bulk fluid, while the lateral flagella are expressed when the polar flagella meet too much resistance to turn. These provide swarming motility on surfaces or in viscous fluids.
Archaeal
The archaellum possessed by some members of domain Archea is superficially similar to the bacterial flagellum; in the 1980s, they were thought to be homologous on the basis of gross morphology and behavior. Both flagella and archaella consist of filaments extending outside the cell, and rotate to propel the cell. Archaeal flagella have a unique structure which lacks a central channel. Similar to bacterial type IV pilins, the archaeal flagellins (archaellins) are made with class 3 signal peptides and they are processed by a type IV prepilin peptidase-like enzyme. The archaellins are typically modified by the addition of N-linked glycans which are necessary for proper assembly and/or function.
Discoveries in the 1990s revealed numerous detailed differences between the archaeal and bacterial flagella. These include:
These differences could mean that the bacterial flagella and archaella could be a classic case of biological analogy, or convergent evolution, rather than homology. However, in comparison to the decades of well-publicized study of bacterial flagella (e.g. by Howard Berg), archaella have only recently begun to garner scientific attention.
Terminology
Aiming to emphasize the distinction between the bacterial flagella and the eukaryotic cilia and flagella, some authors attempted to replace the name of these two eukaryotic structures with "undulipodia" (e.g., all papers by Margulis since the 1970s) or "cilia" for both (e.g., Hülsmann, 1992; Adl et al., 2012; most papers of Cavalier-Smith), preserving "flagella" for the bacterial structure. However, the discriminative usage of the terms "cilia" and "flagella" for eukaryotes adopted in this article is still common (e.g., Andersen et al., 1991; Leadbeater et al., 2000).
Internal structure
A eukaryotic flagellum is a bundle of nine fused pairs of microtubule doublets surrounding two central single microtubules. The so-called "9 + 2" structure is characteristic of the core of the eukaryotic flagellum called an axoneme. At the base of a eukaryotic flagellum is a basal body, "blepharoplast" or kinetosome, which is the microtubule organizing center for flagellar microtubules and is about 500 nanometers long. Basal bodies are structurally identical to centrioles. The flagellum is encased within the cell's plasma membrane, so that the interior of the flagellum is accessible to the cell's cytoplasm.
Besides the axoneme and basal body, relatively constant in morphology, other internal structures of the flagellar apparatus are the transition zone (where the axoneme and basal body meet) and the root system (microtubular or fibrilar structures which extends from the basal bodies into the cytoplasm), more variable and useful as indicators of phylogenetic relationships of eukaryotes. Other structures, more uncommon, are the paraflagellar (or paraxial, paraxonemal) rod, the R fiber, and the S fiber. For surface structures, see below.
Mechanism
Each of the outer 9 doublet microtubules extends a pair of dynein arms (an "inner" and an "outer" arm) to the adjacent microtubule; these produce force through ATP hydrolysis. The flagellar axoneme also contains radial spokes, polypeptide complexes extending from each of the outer nine microtubule doublets towards the central pair, with the "head" of the spoke facing inwards. The radial spoke is thought to be involved in the regulation of flagellar motion, although its exact function and method of action are not yet understood.
Flagella vs cilia
The regular beat patterns of eukaryotic cilia and flagella generate motion on a cellular level. Examples range from the propulsion of single cells such as the swimming of spermatozoa to the transport of fluid along a stationary layer of cells such as in the respiratory tract. Though eukaryotic flagella and motile cilia are ultrastructurally identical, the beating pattern of the two organelles can be different. In the case of flagella, the motion is often planar and wave-like, whereas the motile cilia often perform a more complicated three-dimensional motion with a power and recovery stroke.
Intraflagellar transport
Intraflagellar transport, the process by which axonemal subunits, transmembrane receptors, and other proteins are moved up and down the length of the flagellum, is essential for proper functioning of the flagellum, in both motility and signal transduction.
Evolution and occurrence
Eukaryotic flagella or cilia, probably an ancestral characteristic, are widespread in almost all groups of eukaryotes, as a relatively perennial condition, or as a flagellated life cycle stage (e.g., zoids, gametes, zoospores, which may be produced continually or not).
The first situation is found either in specialized cells of multicellular organisms (e.g., the choanocytes of sponges, or the ciliated epithelia of metazoans), as in ciliates and many eukaryotes with a "flagellate condition" (or "monadoid level of organization", see Flagellata, an artificial group).
Flagellated lifecycle stages are found in many groups, e.g., many green algae (zoospores and male gametes), bryophytes (male gametes), pteridophytes (male gametes), some gymnosperms (cycads and Ginkgo, as male gametes), centric diatoms (male gametes), brown algae (zoospores and gametes), oomycetes (assexual zoospores and gametes), hyphochytrids (zoospores), labyrinthulomycetes (zoospores), some apicomplexans (gametes), some radiolarians (probably gametes), foraminiferans (gametes), plasmodiophoromycetes (zoospores and gametes), myxogastrids (zoospores), metazoans (male gametes), and chytrid fungi (zoospores and gametes).
Flagella or cilia are completely absent in some groups, probably due to a loss rather than being a primitive condition. The loss of cilia occurred in red algae, some green algae (Zygnematophyceae), the gymnosperms except cycads and Ginkgo, angiosperms, pennate diatoms, some apicomplexans, some amoebozoans, in the sperm of some metazoans, and in fungi (except chytrids).
Typology
A number of terms related to flagella or cilia are used to characterize eukaryotes. According to surface structures present, flagella may be:
According to the number of flagella, cells may be (remembering that some authors use "ciliated" instead of "flagellated":
According to the place of insertion of the flagella:
According to the beating pattern:
Other terms related to the flagellar type: