
Clade Reptiliomorpha Scientific name Amniota Higher classification Reptiliomorpha | Clade AmniotaHaeckel, 1866 | |
 | ||
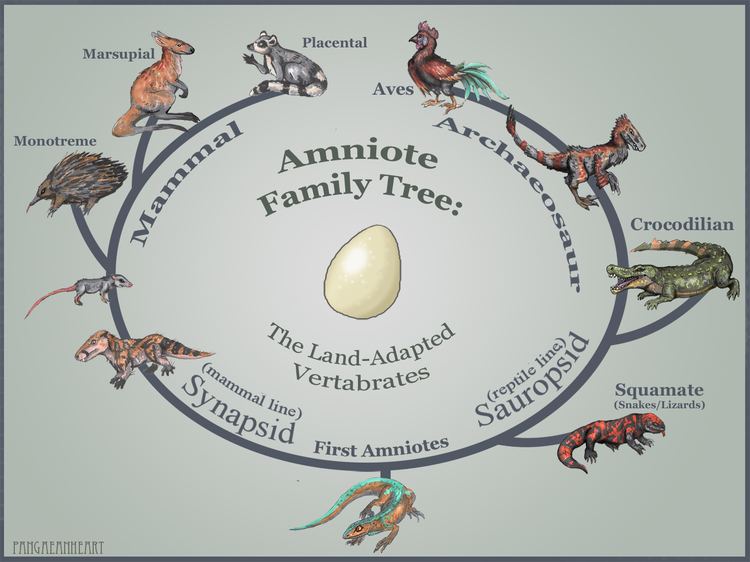
Lower classifications Mammal, Synapsid, Sauropsida, Bear, Macropods | ||
Amniotes
Amniotes (from Greek ἀμνίον amnion, "membrane surrounding the fetus", earlier "bowl in which the blood of sacrificed animals was caught", from ἀμνός amnos, "lamb") are a clade of tetrapod vertebrates comprising the reptiles, birds, and mammals that lay their eggs on land or retain the fertilized egg within the mother. They are distinguished from the anamniotes (fishes and amphibians), which typically lay their eggs in water. Older sources, particularly prior to the 20th century, may refer to amniotes as "higher vertebrates" and anamniotes as "lower vertebrates", based on the discredited idea of the Great Chain of Being.
Contents
- Amniotes
- Cladogram amniotes
- Description
- Adaptions for a terrestrial life
- The egg membranes
- Amniote traits
- Definition and classification
- Traditional classification
- Classification into monophyletic taxa
- Phylogenetic classification
- Cladogram
- References

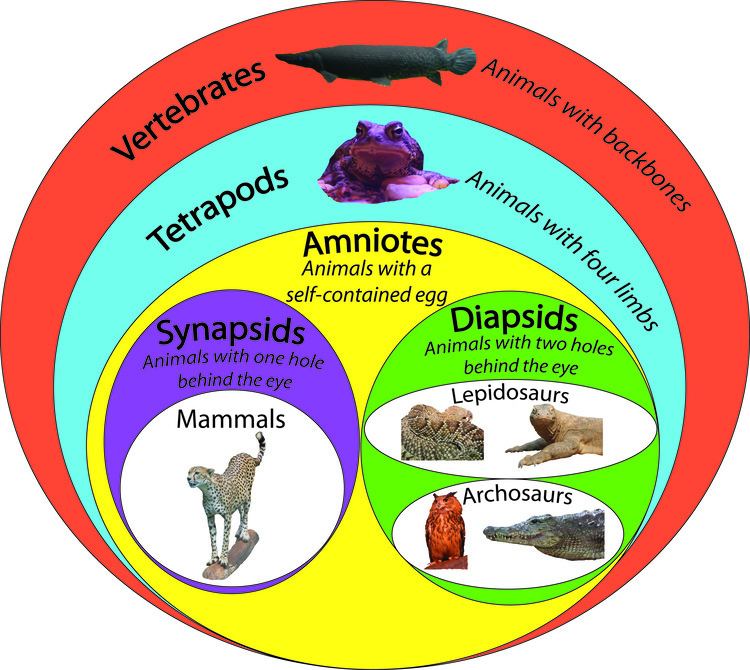
Amniotes are tetrapods (descendants of four-limbed and backboned animals) that are characterised by having an egg equipped with an amnion, an adaptation to lay eggs on land rather than in water as the anamniotes (including frogs) typically do. Amniotes include synapsids (mammals along with their extinct kin) and sauropsids (reptiles and birds), as well as their ancestors, back to amphibians. Amniote embryos, whether laid as eggs or carried by the female, are protected and aided by several extensive membranes. In eutherian mammals (such as humans), these membranes include the amniotic sac that surrounds the fetus. These embryonic membranes and the lack of a larval stage distinguish amniotes from tetrapod amphibians.
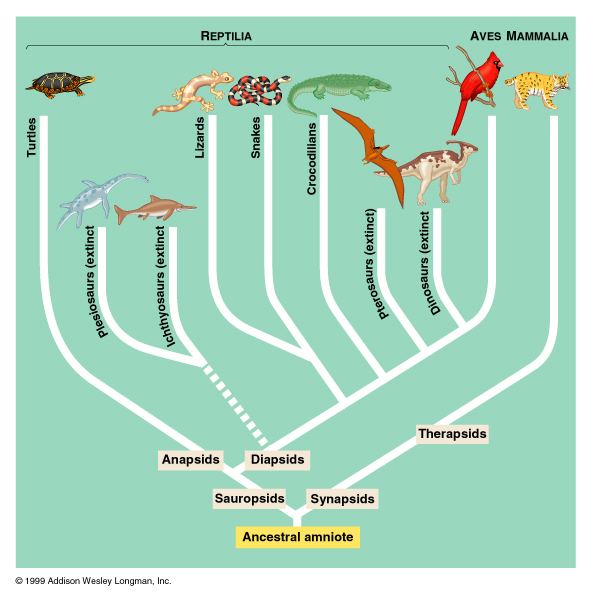
The first amniotes, referred to as "basal amniotes", resembled small lizards and evolved from the amphibian reptiliomorphs about 312 million years ago, in the Carboniferous geologic period. Their eggs could survive out of the water, allowing amniotes to branch out into drier environments. The eggs could also "breathe" and cope with wastes, allowing the eggs and the amniotes themselves to evolve into larger forms.
The amniotic egg represents a critical divergence within the vertebrates, one enabled to reproduce on dry land—free of the need to return to water for reproduction as required of the amphibians. From this point the amniotes spread around the globe, eventually to become the dominant land vertebrates. Very early in the evolutionary history of amniotes, basal amniotes diverged into two main lines, the synapsids and the sauropsids, both of which persist into the modern era. The oldest known fossil synapsid is Protoclepsydrops from about 312 million years ago, while the oldest known sauropsid is probably Paleothyris, in the order Captorhinida, from the Middle Pennsylvanian epoch (ca. 306–312 million years ago).
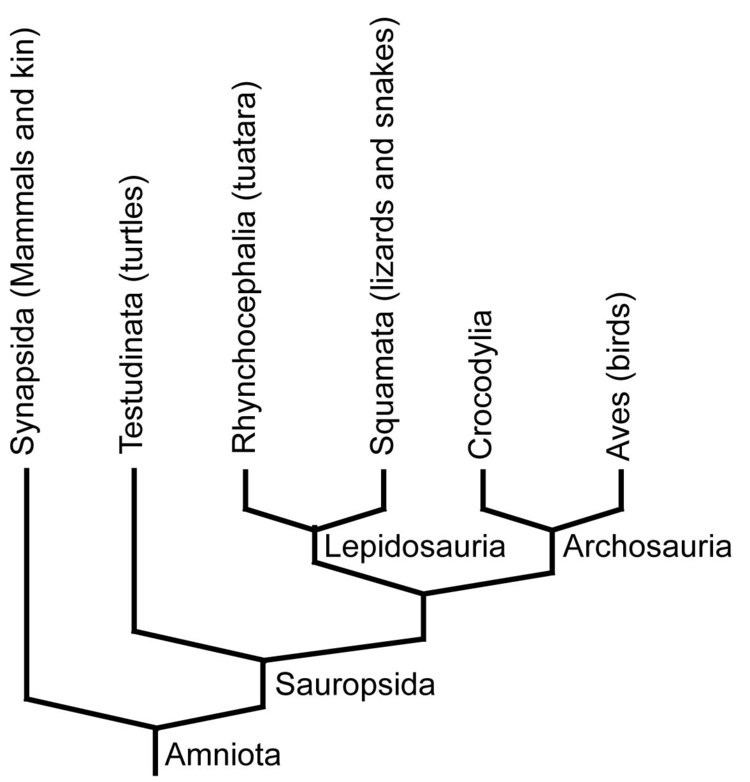
Cladogram amniotes
Description
Amniotes can be characterized in part by embryonic development that includes the formation of several extensive membranes, the amnion, chorion, and allantois. Amniotes develop directly into a (typically) terrestrial form with limbs and a thick stratified epithelium, rather than first entering a feeding larval tadpole stage followed by metamorphosis as in amphibians. In amniotes, the transition from a two-layered periderm to cornified epithelium is triggered by thyroid hormone during embryonic development, rather than metamorphosis. The unique embryonic features of amniotes may reflect specializations of eggs to survive drier environments; or the massive size and yolk content of eggs may have evolved to allow direct development of the embryo to a larger size.
Adaptions for a terrestrial life
Features of amniotes evolved for survival on land include a sturdy but porous leathery or hard eggshell and an allantois evolved to facilitate respiration while providing a reservoir for disposal of wastes. Their kidneys and large intestines are also well-suited to water retention. Most mammals do not lay eggs, but corresponding structures may be found inside the placenta.
The ancestors of true amniotes, such as Casineria kiddi, which lived about 340 million years ago, evolved from amphibian reptiliomorphs and resembled small lizards. At the late Devonian mass extinction 360 million years ago, all known tetrapods were essentially aquatic and fish-like. Because the reptiliomorphs were already established 20 million years later when all their fishlike relatives were extinct, it appears they separated from the other tetrapods somewhere during Romer's gap, when the adult tetrapods became fully terrestrial (some forms would later become secondary aquatic). The modest sized ancestors of the amniotes laid their eggs in moist places, such as depressions under fallen logs or other suitable places in the Carboniferous swamps and forests; and dry conditions were probably not the reason the soft shell emerged. Indeed, many modern day amniotes are dependent on moisture to keep their eggs from desiccating. Although some modern amphibians lay eggs on land, they all lack advanced traits like an amnion. The amniotic egg was formed through a series of evolutionary steps. After internal fertilization and the habit of laying eggs in terrestrial environments became a reproduction strategy amongst the amniote ancestors, the next major breakthrough appears to have been a gradual replacement of the gelatinous coating covering the amphibian egg with a fibrous shell membrane. This allowed the egg to increase both its size and the rate of gas exchange, permitting a larger, metabolically more active embryo to reach full development before hatching. Further development of the design, like extraembryonic membranes (amnion, chorion, and allantois) and a calcified shell, were not essential and probably evolved later. It has been suggested that shelled terrestrial eggs without extraembryonic membranes could still not have been bigger than about 1 cm (0.4 inch) in diameter because of diffusion problems, like the inability to get rid of carbon dioxide if the egg was larger. The only way for the eggs to increase in size would be to develop new internal structures specialized for respiration and waste products. As this happened, it would also affect how much the juveniles could grow before they reached adulthood.
The egg membranes
In fish and amphibians there is only one inner membrane, the embryonic membrane. The amniote egg evolved new internal structures to accommodate gas exchange between the embryo and the atmosphere and to deal with wastes. After these structures developed, further adaption allowed amniotes to lay bigger eggs and in drier habitats. The evolution of larger eggs opened up for larger offspring and consequently larger adults, with those who adapted to a diet of fish and other aquatic animals getting bigger than the terrestrial tetrapods. Further growth for the latter, however, was limited by their position in the terrestrial food chain, which was restricted to level three and below, with only invertebrates occupying level two. Amniotes would eventually experience adaptive radiations when some species evolved the ability to digest plants and new ecological niches opened up, permitting larger body size for both herbivores, omnivores and predators.
Amniote traits
While the early amniotes resembled their amphibian ancestors in many respects, a key difference was the lack of an otic notch at the back margin of the skull roof. In their ancestors, this notch held a spiracle, an unnecessary structure in an animal without an aquatic larval stage. There are three main lines of amniotes, which may be distinguished by the structure of the skull and in particular the number of temporal fenestrae (openings) behind each eye. In anapsids, the ancestral condition, there are none, in synapsids (mammals and their extinct relatives) there is one, and in most diapsids (including birds, crocodilians, squamates, and tuataras), there are two. Turtles were traditionally classified as anapsids because they lack fenestrae, but molecular testing firmly places them in the diapsid line of descent and they therefore secondarily lost their fenestrae.
Post cranial remains of amniotes can be identified from their Labyrinthodont ancestors by their having at least two pairs of sacral ribs, a sternum in the pectoral girdle (some amniotes have lost it) and an astragalus bone in the ankle.
Definition and classification
Amniota was first formally described by embryologist Ernst Haeckel in 1866 on the presence of the amnion, hence the name. A problem with this definition is that the trait (apomorphy) in question does not fossilize, and the status of fossil forms has to be inferred from other traits.
Traditional classification
Classifications of the amniotes have traditionally recognised three classes based on major traits and physiology:
This rather orderly scheme is the one most commonly found in popular and basic scientific works. It has come under critique from cladistics, as the class Reptilia is paraphyletic—it has given rise to two other classes not included in Reptilia.
Classification into monophyletic taxa
A different approach is adopted by writers who reject paraphyletic groupings. One such classification, by Michael Benton, is presented in simplified form below.
Phylogenetic classification
With the advent of cladistics, other researchers have attempted to establish new classes, based on phylogeny, but disregarding the physiological and anatomical unity of the groups. Unlike Benton, for example, Jacques Gauthier and colleagues forwarded a definition of Amniota in 1988 as "the most recent common ancestor of extant mammals and reptiles, and all its descendants". As Gauthier makes use of a crown group definition, Amniota has a slightly different content than the biological amniotes as defined by an apomorphy.
Cladogram
The cladogram presented here illustrates the phylogeny (family tree) of amniotes, and follows a simplified version of the relationships found by Laurin & Reisz (1995). The cladogram covers the group as defined under Gauthier's definition.
The inclusion of Testudines within Parareptilia is unsupported by more recent morphological phylogenetic studies, which placed turtles firmly within diapsids. All molecular studies have also strongly upheld the placement of turtles within diapsids. Within diapsids, some place turtles within Archosauria, or, more commonly, as a sister group to extant archosaurs, though an analysis conducted by Lyson et al. (2012) recovered turtles as the sister group of lepidosaurs instead.