
 | ||
Similar Singlet oxygen, Hydroxide, Hydroxyl radical | ||
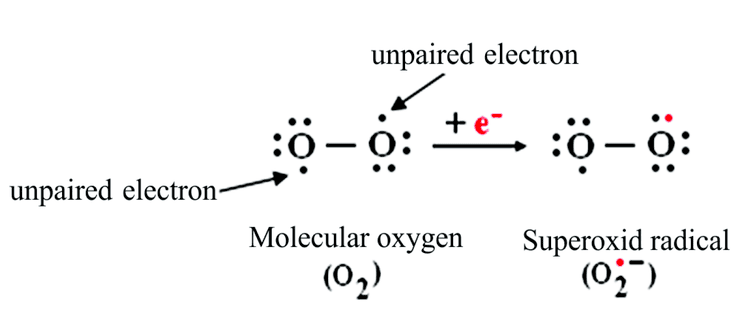
A superoxide, also known by the obsolete name hyperoxide, is a compound that contains the superoxide anion with the Chemical formula O−
2. The systematic name of the anion is dioxide(1−). The reactive oxygen anion superoxide is particularly important as the product of the one-electron reduction of dioxygen O2, which occurs widely in nature. Whereas molecular Oxygen (dioxygen) is a diradical containing two unpaired electrons, the addition of a second electron fills one of its two degenerate molecular orbitals, leaving a charged ionic species with single unpaired electron and a net negative charge of −1. Both dioxygen and the superoxide anion are free radicals that exhibit paramagnetism.
Contents
- Oxide peroxide or superoxide 006
- Properties
- Biology
- FeIII superoxo intermediates
- Assay in biological systems
- References
Oxide peroxide or superoxide 006
Properties
Superoxides are compounds in which the oxidation number of oxygen is − 1⁄2. The O–O bond distance in O−
2 is 1.33 Å, vs. 1.21 Å in O2 and 1.49 Å in O2−
2.
The salts CsO2, RbO2, KO2, and NaO2 are prepared by the direct reaction of O2 with the respective alkali metal. The overall trend corresponds to a reduction in the bond order from 2 (O2), to 1.5 (O−
2), to 1 (O2−
2).

The alkali salts of O−
2 are orange-yellow in color and quite stable, provided they are kept dry. Upon dissolution of these salts in water, however, the dissolved O−
2 undergoes disproportionation (dismutation) extremely rapidly (in a pH-dependent manner):
2 + 2 H2O → 3 O2 + 4 OH−
This reaction (with moisture and carbon dioxide in exhaled air) is the basis of the use of potassium superoxide as an oxygen source in chemical oxygen generators, such as those used on the space shuttle and on submarines. Superoxides are also used in firefighters' oxygen tanks in order to provide a readily available source of oxygen.
In this process O−
2 acts as a Brønsted base, initially forming the radical HO2·. But the pKa of its conjugate acid, hydrogen superoxide (HO2·, also known as "hydroperoxyl" or "perhydroxy radical"), is 4.88 so that at neutral pH 7 all but 0.3% of superoxide is in the anionic form, O−
2.
Potassium superoxide can be dissolved in dimethyl sulfoxide (facilitated by crown ethers) and is stable as long as protons are not available. Superoxide can also be generated in aprotic solvents by cyclic voltammetry.
Salts also decompose in the solid state, but this process requires heating:
2 NaO2 → Na2O2 + O2The derivatives of dioxygen, O2, have characteristic O–O distances that correlate with the bond order of the O–O bond.
Biology
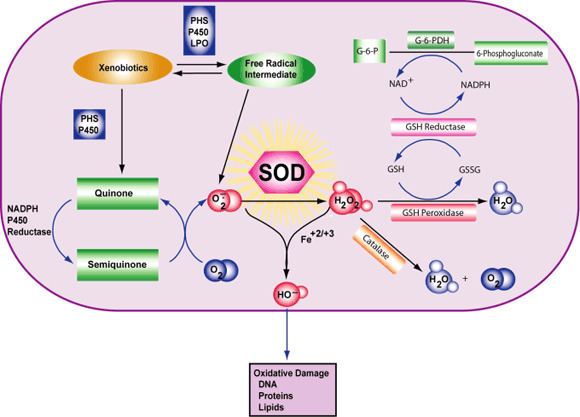
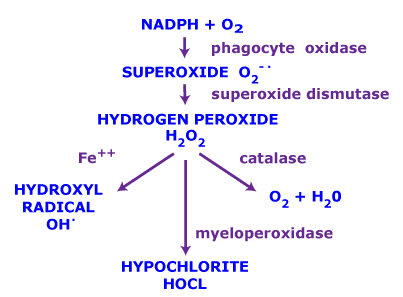
Superoxide is biologically quite toxic and is deployed by the immune system to kill invading microorganisms. In phagocytes, superoxide is produced in large quantities by the enzyme NADPH oxidase for use in oxygen-dependent killing mechanisms of invading pathogens. Mutations in the gene coding for the NADPH oxidase cause an immunodeficiency syndrome called chronic granulomatous disease, characterized by extreme susceptibility to infection, especially catalase-positive organisms. In turn, micro-organisms genetically engineered to lack superoxide dismutase (SOD) lose virulence. Superoxide is also deleterious when produced as a byproduct of mitochondrial respiration (most notably by Complex I and Complex III), as well as several other enzymes, for example xanthine oxidase.
Because superoxide is toxic, nearly all organisms living in the presence of oxygen contain isoforms of the superoxide-scavenging enzyme superoxide dismutase, or SOD. SOD is an extremely efficient enzyme; it catalyzes the neutralization of superoxide nearly as quickly as the two can diffuse together spontaneously in solution. Other proteins that can be both oxidized and reduced by superoxide (e.g., hemoglobin) have weak SOD-like activity. Genetic inactivation ("knockout") of SOD produces deleterious phenotypes in organisms ranging from bacteria to mice and have provided important clues as to the mechanisms of toxicity of superoxide in vivo.
Yeast lacking both mitochondrial and cytosolic SOD grow very poorly in air, but quite well under anaerobic conditions. Absence of cytosolic SOD causes a dramatic increase in mutagenesis and genomic instability. Mice lacking mitochondrial SOD (MnSOD) die around 21 days after birth due to neurodegeneration, cardiomyopathy, and lactic acidosis. Mice lacking cytosolic SOD (CuZnSOD) are viable but suffer from multiple pathologies, including reduced lifespan, liver cancer, muscle atrophy, cataracts, thymic involution, haemolytic anemia and a very rapid age-dependent decline in female fertility.
Superoxide may contribute to the pathogenesis of many diseases (the evidence is particularly strong for radiation poisoning and hyperoxic injury), and perhaps also to aging via the oxidative damage that it inflicts on cells. While the action of superoxide in the pathogenesis of some conditions is strong (for instance, mice and rats overexpressing CuZnSOD or MnSOD are more resistant to strokes and heart attacks), the role of superoxide in aging must be regarded as unproven for now. In model organisms (yeast, the fruit fly Drosophila and mice), genetically knocking out CuZnSOD shortens lifespan and accelerates certain features of aging (cataracts, muscle atrophy, macular degeneration, thymic involution). But the converse, increasing the levels of CuZnSOD, does not seem (except perhaps in Drosophila), to consistently increase lifespan. The most widely accepted view is that oxidative damage (resulting from multiple causes, including superoxide) is but one of several factors limiting lifespan.
Fe(III)-superoxo intermediates
The first step in the activation of O2 by iron-porphyrin complexes involves binding to the Fe(II) ion. Initial attempts to prepare such 1:1 Fe-O2 adducts were met with difficulty due to the propensity of these early models to undergo auto-oxidation via the intermediacy of (μ-η1:η1-peroxo)diiron intermediates. Building upon the early, now classic work in this area, recent studies have provided new insights. The presence of superoxide has been confirmed by various techniques, including L-edge X-ray absorption spectroscopy and Mössbauer spectroscopy.
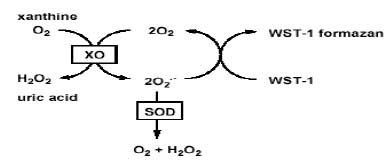
Assay in biological systems
The assay of superoxide generated in biological systems is a difficult task because of its high reactivity and short half-life. One approach that has been used in quantitative assays converts superoxide to hydrogen peroxide, which is relatively stable. Hydrogen Peroxide is then assayed by a fluorimetric method. As a free radical, superoxide has a strong EPR signal, and it is possible to detect superoxide directly using this method when its abundance is high enough. For practical purposes, this can be achieved only in vitro under non-physiological conditions, such as high pH (which slows the spontaneous dismutation) with the enzyme xanthine oxidase. Researchers have developed a series of tool compounds termed "spin traps"(see spin trapping) that can react with superoxide, forming a meta-stable radical (half-life 1–15 minutes), which can be more readily detected by EPR. Superoxide spin trapping was initially carried out with DMPO, but, more recently, phosphorus derivatives with improved half-lives, such as DEPPMPO and DIPPMPO, have become more widely used.