
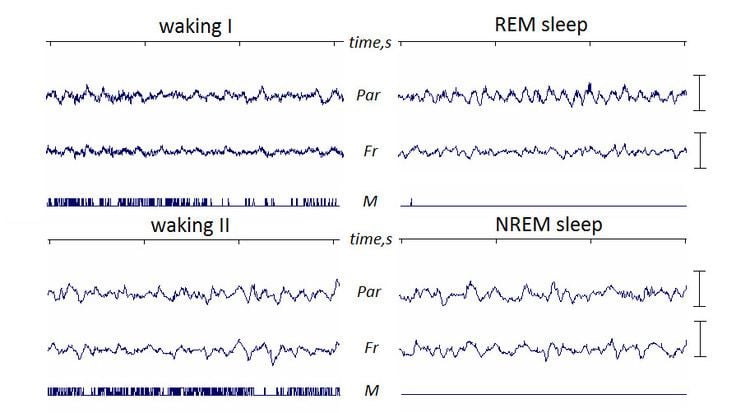
 | ||
Rapid eye movement sleep (REM sleep, REMS) is a unique phase of mammalian sleep characterized by random movement of the eyes, low muscle tone throughout the body, and the propensity of the sleeper to dream vividly. This phase is also known as paradoxical sleep (PS) and sometimes desynchronized sleep because of physiological similarities to waking states, including rapid, low-voltage desynchronized brain waves. Electrical and chemical activity regulating this phase seems to originate in the brain stem and is characterized most notably by an abundance of the neurotransmitter acetylcholine, combined with a nearly complete absence of monoamine neurotransmitters histamine, serotonin, and norepinephrine. The cortical and thalamic neurons of the waking or paradoxically sleeping brain are more depolarized—i.e., can "fire" more readily—than in the deeply sleeping brain. The right and left hemispheres of the brain are more coherent in REM sleep, especially during lucid dreams.
Contents
- Chemicals in brain
- Role of brain stem
- Eye movements
- Circulation respiration and thermoregulation
- Muscle
- Dreaming
- Creativity
- Timing
- Effects of REM sleep deprivation
- REM sleep in animals
- Hypotheses about the functions of REM sleep
- Memory
- Stimulation of the central nervous systems development as a primary function
- Defensive immobilization the precursor of dreams
- Shift of gaze
- Oxygen supply to cornea
- Other theories
- Discovery and further research
- References
REM sleep is punctuated and immediately preceded by PGO (ponto-geniculo-occipital) waves, bursts of electrical activity originating in the brain stem. These waves occur in clusters about every 6 seconds for 1–2 minutes during the transition from deep to paradoxical sleep. They exhibit their highest amplitude upon moving into the visual cortex and are a cause of the "rapid eye movements" in paradoxical sleep.
Brain energy use in REM sleep, as measured by oxygen and glucose metabolism, equals or exceeds energy use in waking. The rate in non-REM sleep is 11–40% lower.
Chemicals in brain
Compared to slow-wave sleep, both waking and paradoxical sleep involve higher use of the neurotransmitter acetylcholine, which may cause the faster brainwaves. The monoamine neurotransmitters norepinephrine, serotonin and histamine are completely unavailable. Injections of acetylcholinesterase inhibitor, which effectively increases available acetylcholine, have been found to induce paradoxical sleep in humans and other animals already in slow-wave sleep. Carbachol, which mimics the effect of acetylcholine on neurons, has a similar influence. In waking humans, the same injections produce paradoxical sleep only if the monoamine neurotransmitters have already been depleted.
Two other neurotransmitters, orexin and gamma-Aminobutyric acid (GABA), seem to promote wakefulness, diminish during deep sleep, and inhibit paradoxical sleep.
Unlike the abrupt transitions in electrical patterns, the chemical changes in the brain show continuous periodic oscillation.
Role of brain stem
Neural activity during REM sleep seems to originate in the brain stem, especially the pontine tegmentum and locus coeruleus. According to the activation-synthesis hypothesis proposed by Robert McCarley and Allan Hobson in 1975–1977, control over REM sleep involves pathways of "REM-on" and "REM-off" neurons in the brain stem. REM-on neurons are primarily cholinergic (i.e., involve acetylcholine); REM-off neurons activate serotonin and noradrenaline, which among other functions suppress the REM-on neurons. McCarley and Hobson suggested that the REM-on neurons actually stimulate REM-off neurons, thereby serving as the mechanism for the cycling between REM and non-REM sleep. They used Lotka–Volterra equations to describe this cyclical inverse relationship. Kayuza Sakai and Michel Jouvet advanced a similar model in 1981. Whereas acetylcholine manifests in the cortex equally during wakefulness and REM, it appears in higher concentrations in the brain stem during REM. The withdrawal of orexin and GABA may cause the absence of the other excitatory neurotransmitters.
Research in the 1990s using positron emission tomography confirmed the role of the brain stem. It also suggested that, within the forebrain, the limbic and paralimbic systems, generally connected with emotion showed more activation than other areas. The areas activated during REM sleep are approximately inverse to those activated during non-REM sleep.
Eye movements
Most of the eye movements in “rapid eye movement” sleep are in fact more rapid than those normally exhibited by waking humans. They are also longer in duration and more likely to loop back to their starting point. About six of such loops take place over one minute of REM sleep. Whereas in slow-wave sleep the eyes can drift apart, the eyes of the paradoxical sleeper move in tandem. These eye movements follow the ponto-geniculo-occipital waves originating in the brain stem. The eye movements themselves may relate to the sense of vision experienced in the dream, but a direct relationship remains to be clearly established. Congenitally blind people, who do not typically have visual imagery in their dreams, still move their eyes in REM sleep. An alternative explanation suggests that the functional purpose of REM sleep is for procedural memory processing, and the rapid eye movement is only an external manifestation of the brain processing the eye-related procedural memory.
Circulation, respiration, and thermoregulation
Generally speaking, the body suspends homeostasis during paradoxical sleep. Heart rate, cardiac pressure, cardiac output, arterial pressure, and breathing rate quickly become irregular when the body moves into REM sleep. In general, respiratory reflexes such as response to hypoxia diminish. Overall, the brain exerts less control over breathing; electrical stimulation of respiration-linked brain areas does not influence the lungs, as it does during non-REM sleep and in waking. The fluctuations of heart rate and arterial pressure tend to coincide with PGO waves and rapid eye movements, twitches, or sudden changes in breathing.
Erections of the penis (nocturnal penile tumescence or NPT) normally accompany REM sleep in rats and humans. If a male has erectile dysfunction (ED) while awake, but has NPT episodes during REM, it would suggest that the ED is from a psychological rather than a physiological cause. In females, erection of the clitoris (nocturnal clitoral tumescence or NCT) causes enlargement, with accompanying vaginal blood flow and transudation (i.e. lubrication). During a normal night of sleep the penis and clitoris may be erect for a total time of from one hour to as long as three and a half hours during REM.
Body temperature is not well regulated during REM sleep, and thus organisms become more sensitive to temperatures outside their thermoneutral zone. Cats and other small furry mammals will shiver and breathe faster to regulate temperature during NREMS but not during REMS. With the loss of muscle tone, animals lose the ability to regulate temperature through body movement. (However, even cats with pontine lesions preventing muscle atonia during REM did not regulate their temperature by shivering.) Neurons which typically activate in response to cold temperatures—triggers for neural thermoregulation—simply do not fire during REM sleep, as they do in NREM sleep and waking.
Consequently, hot or cold environmental temperatures can reduce the proportion of REM sleep, as well as amount of total sleep. In other words, if at the end of a phase of deep sleep, the organism's thermal indicators fall outside of a certain range, it will not enter paradoxical sleep lest deregulation allow temperature to drift further from the desirable value. This mechanism can be 'fooled' by artificially warming the brain.
Muscle
REM atonia, an almost complete paralysis of the body, is accomplished through the inhibition of motor neurons. When the body shifts into REM sleep, motor neurons throughout the body undergo a process called hyperpolarization: their already-negative membrane potential decreases by another 2–10 millivolts, thereby raising the threshold which a stimulus must overcome to excite them. Muscle inhibition may result from unavailability of monoamine neurotransmitters (restraining the abundance of acetylcholine in the brainstem) and perhaps from mechanisms used in waking muscle inhibition. The medulla oblongata, located between pons and spine, seems to have the capacity for organism-wide muscle inhibition. Some localized twitching and reflexes can still occur.
Lack of REM atonia causes REM behavior disorder, sufferers of which physically act out their dreams. (An alternative explanation of this relationship is that the sleeper "dreams out the act": that the muscle impulse precedes the mental image. This explanation could also apply to normal sleepers whose commands to their muscles are suppressed.) (Note that conventional sleepwalking takes place during slow-wave sleep.) Narcolepsy by contrast seems to involve excessive and unwanted REM atonia—i.e., cataplexy and excessive daytime sleepiness while awake, hypnagogic hallucinations before entering slow-wave sleep, or sleep paralysis while waking. Other psychiatric disorders including depression have been linked to disproportionate REM sleep. Patients with suspected sleep disorders are typically evaluated by polysomnogram.
Lesions of the pons to prevent atonia have induced functional “REM behavior disorder” in animals.
Dreaming
Rapid eye movement sleep (REM) has since its discovery been closely associated with dreaming. Waking up sleepers during a REM phase is a common experimental method for obtaining dream reports; 80% of neurotypical people can give some kind of dream report under these circumstances. Sleepers awakened from REM tend to give longer more narrative descriptions of the dreams they were experiencing, and to estimate the duration of their dreams as longer. Lucid dreams are reported far more often in REM sleep. (In fact these could be considered a hybrid state combining essential elements of REM sleep and waking consciousness.) The mental events which occur during REM most commonly have dream hallmarks including narrative structure, convincingness (experiential resemblance to waking life), and incorporation of instinctual themes.
Hobson and McCarley proposed that the PGO waves characteristic of “phasic” REM might supply the visual cortex and forebrain with electrical excitement which amplifies the hallucinatory aspects of dreaming. However, people woken up during sleep do not report significantly more bizarre dreams during phasic REMS, compared to tonic REMS. Another possible relationship between the two phenomena could be that the higher threshold for sensory interruption during REM sleep allows the brain to travel further along unrealistic and peculiar trains of thought.
Some dreaming can take place during non-REM sleep. “Light sleepers” can experience dreaming during stage 2 non-REM sleep, whereas “deep sleepers”, upon awakening in the same stage, are more likely to report “thinking” but not “dreaming”. Certain scientific efforts to assess the uniquely bizarre nature of dreams experienced while asleep were forced to conclude that waking thought could be just as bizarre, especially in conditions of sensory deprivation. Because of non-REM dreaming, some sleep researchers have strenuously contested the importance of connecting dreaming to the REM sleep phase. The prospect that well-known neurological aspects of REM do not themselves cause dreaming suggests the need to re-examine the neurobiology of dreaming per se. Some researchers (Dement, Hobson, Jouvet, for example) tend to resist the idea of disconnecting dreaming from REM sleep.
Creativity
After waking from REM sleep, the mind seems “hyperassociative”—more receptive to semantic priming effects. People awakened from REM have performed better on tasks like anagrams and creative problem solving.
Sleep aids the process by which creativity forms associative elements into new combinations that are useful or meet some requirement. This occurs in REM sleep rather than in NREM sleep. Rather than being due to memory processes, this has been attributed to changes during REM sleep in cholinergic and noradrenergic neuromodulation. High levels of acetylcholine in the hippocampus suppress feedback from hippocampus to the neocortex, while lower levels of acetylcholine and norepinephrine in the neocortex encourage the uncontrolled spread of associational activity within neocortical areas. This is in contrast to waking consciousness, where higher levels of norepinephrine and acetylcholine inhibit recurrent connections in the neocortex. REM sleep through this process adds creativity by allowing "neocortical structures to reorganise associative hierarchies, in which information from the hippocampus would be reinterpreted in relation to previous semantic representations or nodes."
Timing
In the ultradian sleep cycle an organism alternates between deep sleep (slow, large, synchronized brain waves) and paradoxical sleep (faster, desynchronized waves). Sleep happens in the context of the larger circadian rhythm, which influences sleepiness and physiological factors based on timekeepers within the body. Sleep can be distributed throughout the day or clustered during one part of the rhythm: in nocturnal animals, during the day, and in diurnal animals, at night. The organism returns to homeostatic regulation almost immediately after REM sleep ends.
During a night of sleep, humans usually experience about four or five periods of REM sleep; they are quite short at the beginning of the night and longer toward the end. Many animals and some people tend to wake, or experience a period of very light sleep, for a short time immediately after a bout of REM. The relative amount of REM sleep varies considerably with age. A newborn baby spends more than 80% of total sleep time in REM. During REM, the activity of the brain's neurons is quite similar to that during waking hours; for this reason, the REM-sleep stage may be called paradoxical sleep.
REM sleep typically occupies 20–25% of total sleep in adult humans: about 90–120 minutes of a night's sleep. The first REM episode occurs about 70 minutes after falling asleep. Cycles of about 90 minutes each follow, with each cycle including a larger proportion of REM sleep.
Infants spend more time in higher REM sleep than adults. The proportion of REM sleep then decreases significantly in childhood. Older people tend to sleep less overall but sleep in REM for about the same absolute time, and therefore spend a greater proportion of sleep in REM.
Rapid eye movement sleep can be subclassified into tonic and phasic modes. Tonic REM is characterized by theta rhythms in the brain; phasic REM is characterized by PGO waves and actual “rapid” eye movements. Processing of external stimuli is heavily inhibited during phasic REM and recent evidence suggests that sleepers are more difficult to arouse from phasic REM than in slow-wave sleep.
Effects of REM sleep deprivation
REM deprivation causes a significant increase in the number of attempts to go into REM stage while asleep. On recovery nights, an individual will most likely move to stage 3 and REM sleep more quickly and experience an REM rebound, which refers to a great increase in the time spent in REM stage over normal levels. These findings are consistent with the idea that REM sleep is biologically necessary.
After the deprivation is complete, mild psychological disturbances, such as anxiety, irritability, hallucinations, and difficulty concentrating may develop and appetite may increase. There are also positive consequences of REM deprivation. Some symptoms of depression are found to be suppressed by REM deprivation; aggression, and eating behavior may increase. Higher noradrenaline is a possible cause of these results. Whether and how long-term REM deprivation has psychological effects remains a matter of controversy. Several reports have indicated that REM deprivation increases aggressive and sexual behavior in laboratory test animals.
It has been suggested that acute REM sleep deprivation can improve certain types of depression when depression appears to be related to an imbalance of certain neurotransmitters. Although sleep deprivation in general annoys most of the population, it has repeatedly been shown to alleviate depression, albeit temporarily. More than half the individuals who experience this relief report it to be rendered ineffective after sleeping the following night. Thus, researchers have devised methods such as altering the sleep schedule for a span of days following a REM deprivation period and combining sleep-schedule alterations with pharmacotherapy to prolong this effect. Though most antidepressants selectively inhibit REM sleep due to their action on monoamines, this effect decreases after long-term use. Sleep deprivation stimulates hippocampal neurogenesis much the same as antidepressants, but whether this effect is driven by REM sleep in particular is unknown.
Animal studies of REM deprivation are markedly different from human studies. There is evidence that REM sleep deprivation in animals has more serious consequences than in humans. This may be because the length of time animals have been REM deprived for is much longer (up to seventy days) or because the various experimental protocols used have been more uncomfortable and painful than those for humans. The “flower pot” method involves placing a laboratory animal above water on a platform so small that it falls off upon losing muscle tone. The naturally rude awakening which results may elicit changes in the organism which necessarily exceed the simple absence of a sleep phase. Another method involves computer monitoring of brain waves, complete with automatic mechanized shaking of the cage when the test animal drifts into REM sleep.
Evidence suggests that REM deprivation in rats impairs learning of new material, but does not affect existing memory. In one study, rats did not learn to avoid a painful stimulus after REM deprivation as well as they could before the deprivation. No learning impairments have been found in humans undergoing one night of REM deprivation. REM deprivation in rats produces an increase in attempts to enter REM, and after deprivation, REM rebound. In rats, as well as cats, REM sleep deprivation increased brain excitability (e.g. electrical amplification of sensory signals), and which lowered the threshold for waking seizures threshold. This increase in brain excitability seems to be similar in humans. One study also found a decrease in hindbrain sensory excitability. The hindbrain was less receptive overall to information in the afferent pathway, because of the increase in the amplification of those pathways that it is receptive to.
REM sleep in animals
REM sleep occurs in all land mammals as well as in birds. Amount of REM sleep and cycling time vary among animals; predators enjoy more REM sleep than prey. Larger animals also tend to stay in REM for longer, possibly because higher thermal inertia of their brains and bodies allows them to tolerate longer suspension of thermoregulation. The period (full cycle of REM and non-REM) lasts for about 90 minutes in humans, 22 minutes in cats, and 12 minutes in rats.
In utero, mammals spend more than half (50–80%) of a 24-hour day in REM sleep.
Mammals and birds were long thought to be the only animals to experience REM sleep. In 2016 however, researchers found that REM sleep could also be observed in lizards. This suggest that REM sleep has a very ancient evolutionary origin and have probably evolved in the common ancestor of all amniotes.
Hypotheses about the function(s) of REM sleep
While the function of REM sleep is not well understood, several theories have been proposed.
Memory
Sleep in general seems to aid memory. REM sleep may favor the preservation of certain types of memories: specifically, procedural memory, spatial memory, and emotional memory. REM sleep seems to increase following intensive learning in rats, especially several hours after, and sometimes for multiple nights after. Experimental REM deprivation has sometimes inhibited memory consolidation, especially regarding complex processes (e.g., how to escape from an elaborate maze). In humans, the best evidence for REM's improvement of memory pertains to learning of procedures—new ways of moving the body (such as trampoline jumping), and new techniques of problem solving. REM deprivation seemed to impair declarative (i.e., factual) memory only in more complex cases, such as memories of longer stories. REM sleep apparently counteracts attempts to suppress certain thoughts.
According to the dual-process hypothesis of sleep and memory, the two major phases of sleep correspond to different types of memory. “Night half” studies have tested this hypothesis with memory tasks either begun before sleep and assessed in the middle of the night, or begun in the middle of the night and assessed in the morning. Slow-wave sleep, part of non-REM sleep, appears to be important for declarative memory. Artificial enhancement of the non-REM sleep improves the next-day recall of memorized pairs of words. Tucker et al. demonstrated that a daytime nap containing solely non-REM sleep enhances declarative memory but not procedural memory. According to the sequential hypothesis the two types of sleep work together to consolidate memory.
Monoamine oxidase (MAO) inhibitors and tricyclic antidepressants can suppress REM sleep and these drugs show no evidence of impairing memory. Some studies show MAO inhibitors improve memory. Moreover, one case study of an individual who had little or no REM sleep due to a shrapnel injury to the brainstem did not find the individual's memory to be impaired. (For a more detailed critique on the link between sleep and memory, see Ref.))
Intimately related to views on REM function in memory consolidation, Graeme Mitchison and Francis Crick have proposed in 1983 that by virtue of its inherent spontaneous activity, the function of REM sleep "is to remove certain undesirable modes of interaction in networks of cells in the cerebral cortex", which process they characterize as "unlearning". As a result, those memories which are relevant (whose underlying neuronal substrate is strong enough to withstand such spontaneous, chaotic activation), are further strengthened, whilst weaker, transient, "noise" memory traces disintegrate. Memory consolidation during paradoxical is specifically correlated with the periods of rapid eye movement, which do not occur continuously. One explanation for this correlation is that the PGO electrical waves, which precede the eye movements, also influence memory. REM sleep could provide a unique opportunity for “unlearning” to occur in basic neural networks involved in homeostasis, which are protected from this “synaptic downscaling” effect during deep sleep.
Stimulation of the central nervous system's development as a primary function
According to another theory, known as the Ontogenetic Hypothesis of REM sleep, this sleep stage (also known as active sleep in neonates) is particularly important to the developing brain, possibly because it provides the neural stimulation that newborns need to form mature neural connections and for proper nervous system development. Studies investigating the effects of active sleep deprivation have shown that deprivation early in life can result in behavioral problems, permanent sleep disruption, decreased brain mass, and result in an abnormal amount of neuronal cell death. Further supporting this theory is the fact that the amount of REM sleep in humans decreases with age, as well as data from other species (see below).
One important theoretical consequence of the Ontogenetic Hypothesis is that REM sleep may have no essentially vital function in the mature brain, i.e., once the development of the central nervous system has completed. However, because processes of neuronal plasticity do not cease altogether in the brain, REM sleep may continue to be implicated in neurogenesis in adults as a source of sustained spontaneous stimulation.
Defensive immobilization: the precursor of dreams
According to Tsoukalas (2012) REM sleep is an evolutionary transformation of a well-known defensive mechanism, the tonic immobility reflex. This reflex, also known as animal hypnosis or death feigning, functions as the last line of defense against an attacking predator and consists of the total immobilization of the animal so that it appears dead. Tsoukalas argues that the neurophysiology and phenomenology of this reaction shows striking similarities to REM sleep; for example, both reactions exhibit brainstem control, paralysis, hypocampal theta rhythm, and thermoregulatory changes.
Shift of gaze
According to "scanning hypothesis", the directional properties of REM sleep are related to a shift of gaze in dream imagery. Against this hypothesis is that such eye movements occur in those born blind and in fetuses in spite of lack of vision. Also, binocular REMs are non-conjugated (i.e., the two eyes do not point in the same direction at a time) and so lack a fixation point. In support of this theory, research finds that in goal-oriented dreams, eye gaze is directed towards the dream action, determined from correlations in the eye and body movements of REM sleep behavior disorder patients who enact their dreams.
Oxygen supply to cornea
Dr. David M. Maurice (1922-2002), an eye specialist and semi-retired adjunct professor at Columbia University, proposed that REM sleep was associated with oxygen supply to cornea when the animal was sleeping thus aqueous humor, the liquid between cornea and iris, was stagnant if not stirred ("stagnant aqueous humor hypothesis"). Among the supportive evidences, he calculated that if aqueous humor was stagnant, oxygen from iris had to reach cornea by diffusion through aqueous humor, which was not sufficient. According to the theory, when the animal is awake, eye movement and/or cool environmental temperature make sure the aqueous humor is able to circulate. When the animal is sleeping, REM provides the much needed stir to aqueous humor. This theory is consistent with the observation that fetuses, as well as eye-sealed newborn animals, spend much time in REM sleep, and that during a normal sleep, a person's REM sleep episodes become progressively longer deeper into the night. However, owls have REM sleep (telling from EEG recording) but during REM sleep owls do not move their head more than Non-REM sleep. This observation can be derived from Figure S1 of a recently published research and it is well known that owls' eyes are nearly immobile.
Other theories
Another theory suggests that monoamine shutdown is required so that the monoamine receptors in the brain can recover to regain full sensitivity. Indeed, if REM sleep is repeatedly interrupted, the person will compensate for it with longer REM sleep, "rebound sleep", at the next opportunity.
Some researchers argue that the perpetuation of a complex brain process such as REM sleep indicates that it serves an important function for the survival of mammalian and avian species. It fulfills important physiological needs vital for survival to the extent that prolonged REM sleep deprivation leads to death in experimental animals. In both humans and experimental animals, REM sleep loss leads to several behavioral and physiological abnormalities. Loss of REM sleep has been noticed during various natural and experimental infections. Survivability of the experimental animals decreases when REM sleep is totally attenuated during infection; this leads to the possibility that the quality and quantity of REM sleep is generally essential for normal body physiology.
The sentinel hypothesis of REM sleep was put forward by Frederick Snyder in 1966. It is based upon the observation that REM sleep in several mammals (the rat, the hedgehog, the rabbit, and the rhesus monkey) is followed by a brief awakening. This does not occur for either cats or humans, although humans are more likely to wake from REM sleep than from NREM sleep. Snyder hypothesized that REM sleep activates an animal periodically, to scan the environment for possible predators. This hypothesis does not explain the muscle paralysis of REM sleep; however, a logical analysis might suggest that the muscle paralysis exists to prevent the animal from fully waking up unnecessarily, and allowing it to return easily to deeper sleep.
Jim Horne, a sleep researcher at Loughborough University, has suggested that REM in modern humans compensates for the reduced need for wakeful food foraging.
Other theories are that they lubricate the cornea, warm the brain, stimulate and stabilize the neural circuits that have not been activated during waking, create internal stimulation to aid development of the CNS, or lack any purpose, being random creations of brain activation.
Discovery and further research
The German scientist Richard Klaue in 1937 first discovered a period of fast electrical activity in the brains of sleeping cats. In 1944, Ohlmeyer reported 90-minute ultradian sleep cycles involving male erections lasting for 25 minutes. At University of Chicago in 1952, Eugene Aserinsky, Nathaniel Kleitman, and William C. Dement, discovered phases of rapid eye movement during sleep, and connected these to dreaming. Their article was published September 10, 1953.
William Dement advanced the study of REM deprivation, with experiments in which subjects were awoken every time their EEG indicated the beginning of REM sleep. He published "The Effect of Dream Deprivation" in June 1960. ("REM deprivation" has become the more common term following subsequent research indicating the possibility of non-REM dreaming.)
Neurosurgical experiments by Michel Jouvet and others in the following two decades added an understanding of atonia and suggested the importance of the pontine tegmentum (dorsolateral pons) in enabling and regulating paradoxical sleep. Jouvet and others found that damaging the reticular formation of the brainstem inhibited this type of sleep. Jouvet coined the name “paradoxical sleep” in 1959 and in 1962 published results indicating that it could occur in a cat with its entire forebrain removed.