
Order Unassigned Rank Family | Scientific name Orthomyxoviridae Higher classification Viral Group V | |
 | ||
Lower classifications Influenza A virus, Influenzavirus B, Influenzavirus C, Bourbon virus, Isavirus | ||
The Orthomyxoviruses (ορθός, orthos, Greek for "straight"; μυξα, myxa, Greek for "mucus") are a family of RNA viruses that includes seven genera: Influenza virus A, Influenza virus B, Influenza virus C, Influenza virus D, Isavirus, Thogotovirus and Quaranjavirus. The first three genera contain viruses that cause influenza in vertebrates, including birds (see also avian influenza), humans, and other mammals. Isaviruses infect salmon; the thogotoviruses are arboviruses, infecting vertebrates and invertebrates, such as ticks and mosquitoes.
Contents
- Classification
- ICTV Taxonomy
- Types
- Influenza A
- Influenza B
- Influenza C
- Influenza D
- Morphology
- Genome
- Structure
- Replication cycle
- Viability and disinfection
- Vaccination and prophylaxis
- References
The three genera of Influenza virus, which are identified by antigenic differences in their nucleoprotein and matrix protein, infect vertebrates as follows:
Classification
In a phylogenetic-based taxonomy, the category "RNA virus" includes the category "negative-sense ssRNA virus", which includes the Order "Mononegavirales", and the Family "Orthomyxovirus" (among others). The genera-associated species and serotypes of Orthomyxovirus are shown in the following table.
ICTV Taxonomy
Group: ssRNA(-)
Types
There are three genera of influenza virus: Influenza virus A, Influenza virus B and Influenza virus C. Each genus includes only one species, or type: Influenza A virus, Influenza B virus, and Influenza C virus, respectively. Influenza A and C infect multiple species, while influenza B almost exclusively infects humans.
Influenza A
Influenza A viruses are further classified, based on the viral surface proteins hemagglutinin (HA or H) and neuraminidase (NA or N). Sixteen H subtypes (or serotypes) and nine N subtypes of influenza A virus have been identified.
Further variation exists; thus, specific influenza strain isolates are identified by a standard nomenclature specifying virus type, geographical location where first isolated, sequential number of isolation, year of isolation, and HA and NA subtype.
Examples of the nomenclature are:
- A/Brisbane/59/2007 (H1N1)
- A/Moscow/10/99 (H3N2).
The type A viruses are the most virulent human pathogens among the three influenza types and cause the most severe disease. The serotypes that have been confirmed in humans, ordered by the number of known human pandemic deaths, are:
Influenza B
Influenza B virus is almost exclusively a human pathogen, and is less common than influenza A. The only other animal known to be susceptible to influenza B infection is the seal. This type of influenza mutates at a rate 2–3 times lower than type A and consequently is less genetically diverse, with only one influenza B serotype. As a result of this lack of antigenic diversity, a degree of immunity to influenza B is usually acquired at an early age. However, influenza B mutates enough that lasting immunity is not possible. This reduced rate of antigenic change, combined with its limited host range (inhibiting cross species antigenic shift), ensures that pandemics of influenza B do not occur.
Influenza C
The influenza C virus infects humans and pigs, and can cause severe illness and local epidemics. However, influenza C is less common than the other types and usually seems to cause mild disease in children.
Influenza D
This is a genus the first members of which were first isolated in 2011. This genus appears to be most closely related to Influenza C from which it diverged several hundred years ago. There are at least two strains of this genus in extant. The main hosts appear to be cattle but this virus has been reported from pigs also.
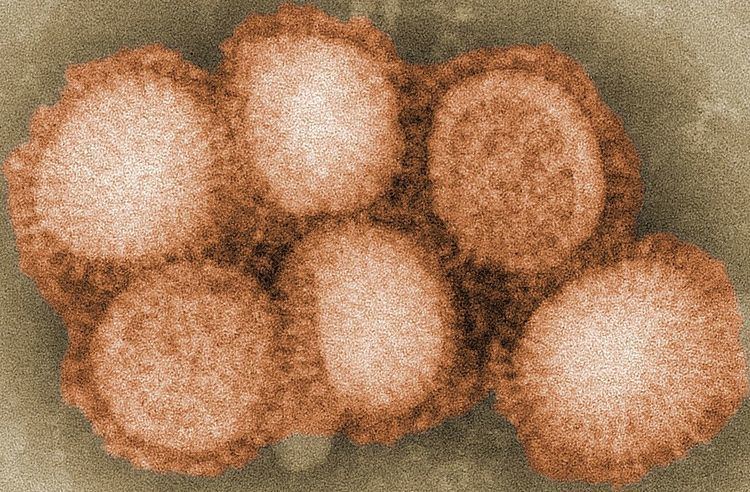
Morphology
The virion is pleomorphic; the envelope can occur in spherical and filamentous forms. In general, the virus's morphology is spherical with particles 50 to 120 nm in diameter, or filamentous virions 20 nm in diameter and 200 to 300 (–3000) nm long. There are some 500 distinct spike-like surface projections of the envelope each projecting 10 to 14 nm from the surface with some types (i.e. hemagglutinin esterase (HEF)) densely dispersed over the surface, and with others (i.e. hemagglutinin (HA)) spaced widely apart.
The major glycoprotein (HA) is interposed irregularly by clusters of neuraminidase (NA), with a ratio of HA to NA of about 4–5 to 1.
Lipoprotein membranes enclose the nucleocapsids; nucleoproteins of different size classes with a loop at each end; the arrangement within the virion is uncertain. The ribonuclear proteins are filamentous and fall in the range of 50 to 130 nm long and 9 to 15 nm in diameter. They have a helical symmetry.
Genome
Viruses of this family contain 6 to 8 segments of linear negative-sense single stranded RNA.
The total genome length is 12000–15000 nucleotides (nt). The largest segment 2300–2500 nt; of second largest 2300–2500 nt; of third 2200–2300 nt; of fourth 1700–1800 nt; of fifth 1500–1600 nt; of sixth 1400–1500 nt; of seventh 1000–1100 nt; of eighth 800–900 nt. Genome sequence has terminal repeated sequences; repeated at both ends. Terminal repeats at the 5'-end 12–13 nucleotides long. Nucleotide sequences of 3'-terminus identical; the same in genera of same family; most on RNA (segments), or on all RNA species. Terminal repeats at the 3'-end 9–11 nucleotides long. Encapsidated nucleic acid is solely genomic. Each virion may contain defective interfering copies.
Structure
The following applies for Influenza A viruses, although other influenza strains are very similar in structure:
The influenza A virus particle or virion is 80–120 nm in diameter and usually roughly spherical, although filamentous forms can occur. Unusually for a virus, the influenza A genome is not a single piece of nucleic acid; instead, it contains eight pieces of segmented negative-sense RNA (13.5 kilobases total), which encode 11 proteins (HA, NA, NP, M1, M2, NS1, NEP, PA, PB1, PB1-F2, PB2). The best-characterised of these viral proteins are hemagglutinin and neuraminidase, two large glycoproteins found on the outside of the viral particles. Neuraminidase is an enzyme involved in the release of progeny virus from infected cells, by cleaving sugars that bind the mature viral particles. By contrast, hemagglutinin is a lectin that mediates binding of the virus to target cells and entry of the viral genome into the target cell. The hemagglutinin (H) and neuraminidase (N) proteins are targets for antiviral drugs. These proteins are also recognised by antibodies, i.e. they are antigens. The responses of antibodies to these proteins are used to classify the different serotypes of influenza A viruses, hence the H and N in H5N1.
Replication cycle
Typically, influenza is transmitted from infected mammals through the air by coughs or sneezes, creating aerosols containing the virus, and from infected birds through their droppings. Influenza can also be transmitted by saliva, nasal secretions, feces and blood. Infections occur through contact with these bodily fluids or with contaminated surfaces. Flu viruses can remain infectious for about one week at human body temperature, over 30 days at 0 °C (32 °F), and indefinitely at very low temperatures (such as lakes in northeast Siberia). They can be inactivated easily by disinfectants and detergents.
The viruses bind to a cell through interactions between its hemagglutinin glycoprotein and sialic acid sugars on the surfaces of epithelial cells in the lung and throat (Stage 1 in infection figure). The cell imports the virus by endocytosis. In the acidic endosome, part of the haemagglutinin protein fuses the viral envelope with the vacuole's membrane, releasing the viral RNA (vRNA) molecules, accessory proteins and RNA-dependent RNA polymerase into the cytoplasm (Stage 2). These proteins and vRNA form a complex that is transported into the cell nucleus, where the RNA-dependent RNA polymerase begins transcribing complementary positive-sense cRNA (Steps 3a and b). The cRNA is either exported into the cytoplasm and translated (step 4), or remains in the nucleus. Newly synthesised viral proteins are either secreted through the Golgi apparatus onto the cell surface (in the case of neuraminidase and hemagglutinin, step 5b) or transported back into the nucleus to bind vRNA and form new viral genome particles (step 5a). Other viral proteins have multiple actions in the host cell, including degrading cellular mRNA and using the released nucleotides for vRNA synthesis and also inhibiting translation of host-cell mRNAs.
Negative-sense vRNAs that form the genomes of future viruses, RNA-dependent RNA transcriptase, and other viral proteins are assembled into a virion. Hemagglutinin and neuraminidase molecules cluster into a bulge in the cell membrane. The vRNA and viral core proteins leave the nucleus and enter this membrane protrusion (step 6). The mature virus buds off from the cell in a sphere of host phospholipid membrane, acquiring hemagglutinin and neuraminidase with this membrane coat (step 7). As before, the viruses adhere to the cell through hemagglutinin; the mature viruses detach once their neuraminidase has cleaved sialic acid residues from the host cell. After the release of new influenza virus, the host cell dies.
Orthomyxoviridae viruses are one of two RNA viruses that replicate in the nucleus (the other being retroviridae). This is because the machinery of orthomyxo viruses cannot make their own mRNAs. They use cellular RNAs as primers for initiating the viral mRNA synthesis in a process known as cap-snatching. Once in the nucleus, the RNA Polymerase Protein PB2 finds a cellular pre-mRNA and binds to its 5' capped end. Then RNA Polymerase PA cleaves off the cellular mRNA near the 5' end and uses this capped fragment as a primer for transcribing the rest of the viral RNA genome in viral mRNA. This is due to the need of mRNA to have a 5' cap in order to be recognized by the cell's ribosome for translation.
Since RNA proofreading enzymes are absent, the RNA-dependent RNA transcriptase makes a single nucleotide insertion error roughly every 10 thousand nucleotides, which is the approximate length of the influenza vRNA. Hence, nearly every newly manufactured influenza virus will contain a mutation in its genome. The separation of the genome into eight separate segments of vRNA allows mixing (reassortment) of the genes if more than one variety of influenza virus has infected the same cell (superinfection). The resulting alteration in the genome segments packaged into viral progeny confers new behavior, sometimes the ability to infect new host species or to overcome protective immunity of host populations to its old genome (in which case it is called an antigenic shift).
Viability and disinfection
Mammalian influenza viruses tend to be labile, but can survive several hours in mucus. Avian influenza virus can survive for 100 days in distilled water at room temperature, and 200 days at 17 °C (63 °F). The avian virus is inactivated more quickly in manure, but can survive for up to 2 weeks in feces on cages. Avian influenza viruses can survive indefinitely when frozen. Influenza viruses are susceptible to bleach, 70% ethanol, aldehydes, oxidizing agents, and quaternary ammonium compounds. They are inactivated by heat of 133 °F (56 °C) for minimum of 60 minutes, as well as by low pH <2.
Vaccination and prophylaxis
Vaccines and drugs are available for the prophylaxis and treatment of influenza virus infections. Vaccines are composed of either inactivated or live attenuated virions of the H1N1 and H3N2 human influenza A viruses, as well as those of influenza B viruses. Because the antigenicities of the wild viruses evolve, vaccines are reformulated annually by updating the seed strains. However, when the antigenicities of the seed strains and wild viruses do not match, vaccines fail to protect the vaccinees. In addition, even when they do match, escape mutants are often generated. Drugs available for the treatment of influenza include Amantadine and Rimantadine, which inhibit the uncoating of virions by interfering with M2, and Oseltamivir (marketed under the brand name Tamiflu), Zanamivir, and Peramivir, which inhibit the release of virions from infected cells by interfering with NA. However, escape mutants are often generated for the former drug and less frequently for the latter drug.