
Group Group V ((−)ssRNA) Higher classification Influenzavirus A | Order Unassigned Genus Influenzavirus A Rank Species | |
 | ||
Lower classifications Influenza A virus subtype, Influenza A virus subtype, Influenza A virus subtype, Influenza A virus subtype, Influenza A virus subtype | ||
H1n1 influenza a virus
Influenza A virus causes influenza in birds and some mammals, and is the only species of influenza virus A. Influenza virus A is a genus of the Orthomyxoviridae family of viruses. Strains of all subtypes of influenza A virus have been isolated from wild birds, although disease is uncommon. Some isolates of influenza A virus cause severe disease both in domestic poultry and, rarely, in humans. Occasionally, viruses are transmitted from wild aquatic birds to domestic poultry, and this may cause an outbreak or give rise to human influenza pandemics.
Contents
- H1n1 influenza a virus
- Variants and subtypes
- Annual flu
- FI6 antibody
- Structure and genetics
- Multiplicity Reactivation
- In Non humans
- Human influenza virus
- Evolution
- References
Influenza A viruses are negative-sense, single-stranded, segmented RNA viruses. The several subtypes are labeled according to an H number (for the type of hemagglutinin) and an N number (for the type of neuraminidase). There are 18 different known H antigens (H1 to H18) and 11 different known N antigens (N1 to N11). H17 was isolated from fruit bats in 2012. H18N11 was discovered in a Peruvian bat in 2013.
Each virus subtype has mutated into a variety of strains with differing pathogenic profiles; some are pathogenic to one species but not others, some are pathogenic to multiple species.
A filtered and purified influenza A vaccine for humans has been developed, and many countries have stockpiled it to allow a quick administration to the population in the event of an avian influenza pandemic. Avian influenza is sometimes called avian flu, and colloquially, bird flu. In 2011, researchers reported the discovery of an antibody effective against all types of the influenza A virus.
Variants and subtypes
Influenza type A viruses are categorized into subtypes based on the type of two proteins on the surface of the viral envelope:
H = hemagglutinin, a protein that causes red blood cells to agglutinate.N = neuraminidase, an enzyme that cleaves the glycosidic bonds of the monosaccharide, neuraminic acidDifferent influenza viruses encode for different hemagglutinin and neuraminidase proteins. For example, the H5N1 virus designates an influenza A subtype that has a type 5 hemagglutinin (H) protein and a type 1 neuraminidase (N) protein. There are 18 known types of hemagglutinin and 11 known types of neuraminidase, so, in theory, 198 different combinations of these proteins are possible.
Some variants are identified and named according to the isolate they resemble, thus are presumed to share lineage (example Fujian flu virus-like); according to their typical host (example human flu virus); according to their subtype (example H3N2); and according to their deadliness (example LP, low pathogenic). So a flu from a virus similar to the isolate A/Fujian/411/2002(H3N2) is called Fujian flu, human flu, and H3N2 flu.
Variants are sometimes named according to the species (host) in which the strain is endemic or to which it is adapted. The main variants named using this convention are:
Variants have also sometimes been named according to their deadliness in poultry, especially chickens:
Most known strains are extinct strains. For example, the annual flu subtype H3N2 no longer contains the strain that caused the Hong Kong flu.
Annual flu
The annual flu (also called "seasonal flu" or "human flu") in the U.S. "results in approximately 36,000 deaths and more than 200,000 hospitalizations each year. In addition to this human toll, influenza is annually responsible for a total cost of over $10 billion in the U.S."
The annually updated, trivalent influenza vaccine consists of hemagglutinin (HA) surface glycoprotein components from influenza H3N2, H1N1, and B influenza viruses.
Measured resistance to the standard antiviral drugs amantadine and rimantadine in H3N2 has increased from 1% in 1994 to 12% in 2003 to 91% in 2005.
"Contemporary human H3N2 influenza viruses are now endemic in pigs in southern China and can reassort with avian H5N1 viruses in this intermediate host."
FI6 antibody
FI6, an antibody that targets the hemagglutinin protein, was discovered in 2011. FI6 is the only known antibody effective against all 16 subtypes of the influenza A virus.
Structure and genetics
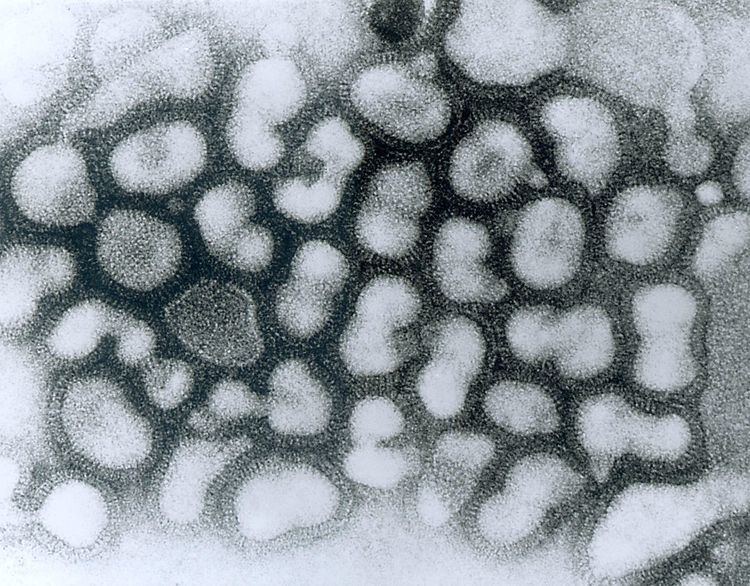
Influenza type A viruses are very similar in structure to influenza viruses types B and C. The virus particle (also called the virion) is 80–120 nanometers in diameter and usually roughly spherical, although some rare filamentous forms can occur. According to researchers, there are more filamentous particles in clinical isolates, whereas laboratory strains consist of more spherical virions.
Despite these varied shapes, the virions of all influenza type A viruses are similar in composition. They are all made up of a viral envelope containing two main types of proteins, wrapped around a central core.
The two large proteins found on the outside of viral particles are hemagglutinin (HA) and neuraminidase (NA). HA is a protein that mediates binding of the virion to target cells and entry of the viral genome into the target cell, while NA is involved in the release of progeny virions from infected cells. These proteins are usually the targets for antiviral drugs. Furthermore, they are also the antigen proteins to which a host’s antibodies can bind and trigger an immune response. Influenza type A viruses are categorized into subtypes based on the type of these two proteins on the surface of the viral envelope. There are 16 subtypes of HA and 9 subtypes of NA known, but only H 1, 2 and 3, and N 1 and 2 are commonly found in humans.
The central core of a virion contains the viral genome and other viral proteins that package and protect the genetic material. Unlike the genomes of most organisms (including humans, animals, plants, and bacteria) which are made up of double-stranded DNA, many viral genomes are made up of a different, single-stranded nucleic acid called RNA. Unusually for a virus, though, the influenza type A virus genome is not a single piece of RNA; instead, it consists of segmented pieces of negative-sense RNA, each piece containing either one or two genes which code for a gene product (protein). The term negative-sense RNA just implies that the RNA genome cannot be translated into protein directly; it must first be transcribed to positive-sense RNA before it can be translated into protein products. The segmented nature of the genome allows for the exchange of entire genes between different viral strains.
The entire Influenza A virus genome is 13,588 bases long and is contained on eight RNA segments that code for 11 proteins:
The RNA segments of the viral genome have complementary base sequences at the terminal ends, allowing them to bond to each other with hydrogen bonds. After transcription from negative-sense to positive-sense RNA takes place, the positive-sense RNA strands are capped on the 5’ end by a process called cap snatching. This involves the viral protein NS1 binding to the host cell’s early messenger RNA transcripts. A second viral protein, PA, cleaves the cap from the host’s RNA. The short cap is then added to the influenza positive-sense RNA strands, allowing it to be processed by ribosomes and translated into its protein products. The positive-sense RNA strands also serve for synthesis of negative-sense RNA strands for new virions.
The RNA synthesis takes place in the cell nucleus, while the synthesis of proteins takes place in the cytoplasm. Once the viral proteins are assembled into virions, the assembled virions leave the nucleus and migrate towards the cell membrane. The host cell membrane has patches of viral transmembrane proteins (HA, NA, and M2) and an underlying layer of the M1 protein which assist the assembled virions to budding through the membrane, releasing finished enveloped viruses into the extracellular fluid.
Multiplicity Reactivation
Influenza virus is able to undergo multiplicity reactivation after inactivation by UV radiation, or by ionizing radiation. If any of the eight RNA strands that make up the genome contains damage that prevents replication or expression of an essential gene, the virus is not viable when it alone infects a cell (a single infection). However, when two or more damaged viruses infect the same cell (multiple infection), viable progeny viruses can be produced provided each of the eight genomic segments is present in at least one undamaged copy. That is, multiplicity reactivation can occur.
Upon infection, influenza virus induces a host response involving increased production of reactive oxygen species, and this can damage the virus genome. If, under natural conditions, virus survival is ordinarily vulnerable to the challenge of oxidative damage, then multiplicity reactivation is likely selectively advantageous as a kind of genomic repair process. It has been suggested that multiplicity reactivation involving segmented RNA genomes may be similar to the earliest evolved form of sexual interaction in the RNA world that likely preceded the DNA world. (Also see RNA world hypothesis.)
In Non-humans
See H5N1 for the current epizootic (an epidemic in nonhumans) and panzootic (a disease affecting animals of many species especially over a wide area) of H5N1 influenzaFowl act as natural asymptomatic carriers of influenza A viruses. Prior to the current H5N1 epizootic, strains of influenza A virus had been demonstrated to be transmitted from wild fowl to only birds, pigs, horses, seals, whales and humans; and only between humans and pigs and between humans and domestic fowl; and not other pathways such as domestic fowl to horse.
Wild aquatic birds are the natural hosts for a large variety of influenza A viruses. Occasionally, viruses are transmitted from these birds to other species and may then cause devastating outbreaks in domestic poultry or give rise to human influenza pandemics.
H5N1 has been shown to be transmitted to tigers, leopards, and domestic cats that were fed uncooked domestic fowl (chickens) with the virus. H3N8 viruses from horses have crossed over and caused outbreaks in dogs. Laboratory mice have been infected successfully with a variety of avian flu genotypes.
Influenza A viruses spread in the air and in manure, and survives longer in cold weather. It can also be transmitted by contaminated feed, water, equipment and clothing; however, there is no evidence the virus can survive in well-cooked meat. Symptoms in animals vary, but virulent strains can cause death within a few days.
"Highly pathogenic avian influenza virus is on every top ten list available for potential agricultural bioweapon agents".
Avian influenza viruses that the OIE and others test for to control poultry disease include: H5N1, H7N2, H1N7, H7N3, H13N6, H5N9, H11N6, H3N8, H9N2, H5N2, H4N8, H10N7, H2N2, H8N4, H14N5, H6N5, H12N5 and others.
*Outbreaks with significant spread to numerous farms, resulting in great economic losses. Most other outbreaks involved little or no spread from the initially infected farms.
1979: "More than 400 harbor seals, most of them immature, died along the New England coast between December 1979 and October 1980 of acute pneumonia associated with influenza virus, A/Seal/Mass/1/180 (H7N7)."
1995: "[V]accinated birds can develop asymptomatic infections that allow virus to spread, mutate, and recombine (ProMED-mail, 2004j). Intensive surveillance is required to detect these “silent epidemics” in time to curtail them. In Mexico, for example, mass vaccination of chickens against epidemic H5N2 influenza in 1995 has had to continue in order to control a persistent and evolving virus (Lee et al., 2004)."
1997: "Influenza A viruses normally seen in one species sometimes can cross over and cause illness in another species. For example, until 1997, only H1N1 viruses circulated widely in the U.S. pig population. However, in 1997, H3N2 viruses from humans were introduced into the pig population and caused widespread disease among pigs. Most recently, H3N8 viruses from horses have crossed over and caused outbreaks in dogs."
2000: "In California, poultry producers kept their knowledge of a recent H6N2 avian influenza outbreak to themselves due to their fear of public rejection of poultry products; meanwhile, the disease spread across the western United States and has since become endemic."
2003: In Netherlands H7N7 influenza virus infection broke out in poultry on several farms.
2004: In North America, the presence of avian influenza strain H7N3 was confirmed at several poultry farms in British Columbia in February 2004. As of April 2004, 18 farms had been quarantined to halt the spread of the virus.
2005: Tens of millions of birds died of H5N1 influenza and hundreds of millions of birds were culled to protect humans from H5N1. H5N1 is endemic in birds in southeast Asia and represents a long-term pandemic threat.
2006: H5N1 spreads across the globe, killing hundreds of millions of birds and over 100 people, and causing a significant H5N1 impact from both actual deaths and predicted possible deaths.
Human influenza virus
"Human influenza virus" usually refers to those subtypes that spread widely among humans. H1N1, H1N2, and H3N2 are the only known influenza A virus subtypes currently circulating among humans.
Genetic factors in distinguishing between "human flu viruses" and "avian influenza viruses" include:
PB2: (RNA polymerase): Amino acid (or residue) position 627 in the PB2 protein encoded by the PB2 RNA gene. Until H5N1, all known avian influenza viruses had a Glu at position 627, while all human influenza viruses had a lysine.HA: (hemagglutinin): Avian influenza HA binds alpha 2–3 sialic acid receptors, while human influenza HA binds alpha 2–6 sialic acid receptors. Swine influenza viruses have the ability to bind both types of sialic acid receptors."About 52 key genetic changes distinguish avian influenza strains from those that spread easily among people, according to researchers in Taiwan, who analyzed the genes of more than 400 A type flu viruses." "How many mutations would make an avian virus capable of infecting humans efficiently, or how many mutations would render an influenza virus a pandemic strain, is difficult to predict. We have examined sequences from the 1918 strain, which is the only pandemic influenza virus that could be entirely derived from avian strains. Of the 52 species-associated positions, 16 have residues typical for human strains; the others remained as avian signatures. The result supports the hypothesis that the 1918 pandemic virus is more closely related to the avian influenza A virus than are other human influenza viruses."
Human flu symptoms usually include fever, cough, sore throat, muscle aches, conjunctivitis and, in severe cases, severe breathing problems and pneumonia that may be fatal. The severity of the infection will depend in large part on the state of the infected person's immune system and if the victim has been exposed to the strain before, and is therefore partially immune.
Highly pathogenic H5N1 avian influenza in a human is far worse, killing 50% of humans who catch it. In one case, a boy with H5N1 experienced diarrhea followed rapidly by a coma without developing respiratory or flu-like symptoms.
The influenza A virus subtypes that have been confirmed in humans, ordered by the number of known human pandemic deaths, are:
Evolution
Taubenberger says:
"All influenza A pandemics since [the Spanish flu pandemic], and indeed almost all cases of influenza A worldwide (excepting human infections from avian viruses such as H5N1 and H7N7), have been caused by descendants of the 1918 virus, including "drifted" H1N1 viruses and reassorted H2N2 and H3N2 viruses. The latter are composed of key genes from the 1918 virus, updated by subsequently incorporated avian influenza genes that code for novel surface proteins, making the 1918 virus indeed the "mother" of all pandemics."Researchers from the National Institutes of Health used data from the Influenza Genome Sequencing Project and concluded that during the ten-year period examined, most of the time the hemagglutinin gene in H3N2 showed no significant excess of mutations in the antigenic regions while an increasing variety of strains accumulated. This resulted in one of the variants eventually achieving higher fitness, becoming dominant, and in a brief interval of rapid evolution, rapidly sweeping through the population and eliminating most other variants.
Short-term Evolution In the short-term evolution of influenza A virus, a 2006 study found that stochastic, or random, processes are key factors. Influenza A virus HA antigenic evolution appears to be characterized more by punctuated, sporadic jumps as opposed to a constant rate of antigenic change. Using phylogenetic analysis of 413 complete genomes of human influenza A viruses that were collected throughout the state of New York, the authors of Nelson et al. 2006 were able to show that genetic diversity, and not antigenic drift, shaped the short-term evolution of influenza A via random migration and reassortment. The evolution of these viruses is dominated more by the random importation of genetically different viral strains from other geographic locations and less by natural selection. Within a given season, adaptive evolution is infrequent and had an overall weak effect as evidenced from the data gathered from the 413 genomes. Phylogenetic analysis revealed the different strains were derived from newly imported genetic material as opposed to isolates that had been circulating in New York in previous seasons. Therefore, the gene flow in and out of this population, and not natural selection, was more important in the short term.