
Order Diptera Infraorder Culicomorpha Scientific name Culicidae Clutch size 100 – 200 | Phylum Arthropoda Suborder Nematocera Higher classification Culicoidea | |
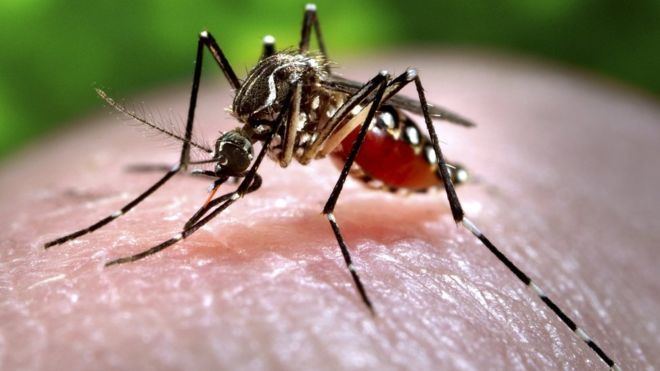 | ||
Lifespan Female: 42 – 56 days (Adult), Male: 10 days (Adult) Lower classifications Aedes, Marsh Mosquitoes, yellow fever mosquito, Asian tiger mosquito, Mansonia | ||
Mosquitoes are small, midge-like flies that constitute the family Culicidae. Females of most species are ectoparasites, whose tube-like mouthparts (called a proboscis) pierce the hosts' skin to consume blood. The word "mosquito" (formed by mosca and diminutive -ito) is Spanish for "little fly". Thousands of species feed on the blood of various kinds of hosts, mainly vertebrates, including mammals, birds, reptiles, amphibians, and even some kinds of fish. Some mosquitoes also attack invertebrates, mainly other arthropods. Though the loss of blood is seldom of any importance to the victim, the saliva of the mosquito often causes an irritating rash that is a serious nuisance. Much more serious though, are the roles of many species of mosquitoes as vectors of diseases. In passing from host to host, some transmit extremely harmful infections such as malaria, yellow fever, Chikungunya, West Nile virus, dengue fever, filariasis, Zika virus and other arboviruses, rendering it the deadliest animal family in the world.
Contents
- How mosquitoes use six needles to suck your blood deep look
- Taxonomy and evolution
- Subfamilies
- Species
- Lifecycle
- Eggs and oviposition
- Larva
- Pupa
- Adult
- Feeding by adults
- Hosts of blood feeding mosquito species
- Mouthparts
- Saliva
- Egg development and blood digestion
- Distribution
- Means of dispersal
- Disease
- Control
- Repellents
- Bites and treatment
- In human culture
- References
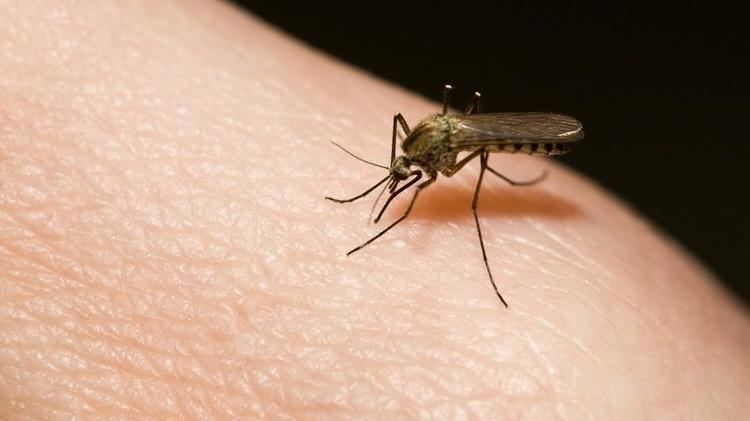
How mosquitoes use six needles to suck your blood deep look
Taxonomy and evolution
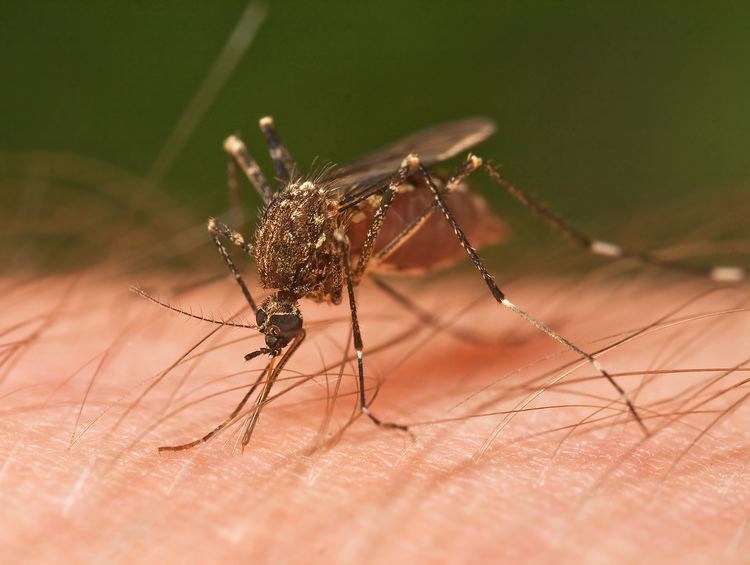
The oldest known mosquito with an anatomy similar to modern species was found in 79-million-year-old Canadian amber from the Cretaceous. An older sister species with more primitive features was found in Burmese amber that is 90 to 100 million years old. Two mosquito fossils have been found that show very little morphological change in modern mosquitoes against their counterpart from 46 million years ago. These fossils are also the oldest ever found to have blood preserved within their abdomens. Despite no fossils being found earlier than the Cretaceous, recent studies suggest that the earliest divergence of mosquitoes between the lineages leading to Anophelinae and Culicinae occurred 226 million years ago.
The Old and New World Anopheles species are believed to have subsequently diverged about 95 million years ago.

The mosquito Anopheles gambiae is currently undergoing speciation into the M(opti) and S(avanah) molecular forms. Consequently, some pesticides that work on the M form no longer work on the S form. Over 3,500 species of the Culicidae have already been described. They are generally divided into two subfamilies which in turn comprise some 43 genera. These figures are subject to continual change, as more species are discovered, and as DNA studies compel rearrangement of the taxonomy of the family. The two main subfamilies are the Anophelinae and Culicinae, with their genera as shown in the subsection below. The distinction is of great practical importance because the two subfamilies tend to differ in their significance as vectors of different classes of diseases. Roughly speaking, arboviral diseases such as yellow fever and dengue fever tend to be transmitted by Culicine species, not necessarily in the genus Culex. Some transmit various species of avian malaria, but it is not clear that they ever transmit any form of human malaria. Some species do however transmit various forms of filariasis, much as many Simuliidae do.

Anopheline mosquitoes, again not necessarily in the genus Anopheles, sometimes bear pathogenic arboviruses, but it is not yet clear that they ever transmit them as effective vectors. However, all the most important vectors of human malaria are Anopheline.
Subfamilies
Species

Mosquitoes are members of a family of nematocerid flies: the Culicidae (from the Latin Culex, genitive culicis, meaning "midge" or "gnat"). Superficially, mosquitoes resemble crane flies (family Tipulidae) and chironomid flies (family Chironomidae). In particular, the females of many species of mosquitoes are blood-eating pests and dangerous vectors of diseases, whereas members of the similar-looking Chironomidae and Tipulidae are not. Many species of mosquitoes are not blood eaters and of those that are, many create a "high to low pressure" in the blood to obtain it and do not transmit disease. Also, in the bloodsucking species, only the females suck blood. Furthermore, even among mosquitoes that do carry important diseases, neither all species of mosquitoes, nor all strains of a given species transmit the same kinds of diseases, nor do they all transmit the diseases under the same circumstances; their habits differ. For example, some species attack people in houses, and others prefer to attack people walking in forests. Accordingly, in managing public health, knowing which species or even which strain of mosquito one is dealing with is important.

Over 3,500 species of mosquitoes have already been described from various parts of the world. Some mosquitoes that bite humans routinely act as vectors for a number of infectious diseases affecting millions of people per year. Others that do not routinely bite humans, but are the vectors for animal diseases, may become disastrous agents for zoonosis of new diseases when their habitats are disturbed, for instance by sudden deforestation.
Lifecycle
Like all flies, mosquitoes go through four stages in their lifecycles: egg, larva, pupa, and adult or imago. In most species, adult females lay their eggs in stagnant water; some lay eggs near the water's edge; others attach their eggs to aquatic plants. Each species selects the situation of the water into which it lays its eggs and does so according to its own ecological adaptations. Some are generalists and are not very fussy. Some breed in lakes, some in temporary puddles. Some breed in marshes, some in salt-marshes. Among those that breed in salt water, some are equally at home in fresh and salt water up to about one-third the concentration of seawater, whereas others must acclimatize themselves to the salinity. Such differences are important because certain ecological preferences keep mosquitoes away from most humans, whereas other preferences bring them right into houses at night.
Some species of mosquitoes prefer to breed in phytotelmata (natural reservoirs on plants), such as rainwater accumulated in holes in tree trunks, or in the leaf-axils of bromeliads. Some specialize in the liquid in pitchers of particular species of pitcher plants, their larvae feeding on decaying insects that had drowned there or on the associated bacteria; the genus Wyeomyia provides such examples — the harmless Wyeomyia smithii breeds only in the pitchers of Sarracenia purpurea.
However, some of the species of mosquitoes that are adapted to breeding in phytotelmata are dangerous disease vectors. In nature, they might occupy anything from a hollow tree trunk to a cupped leaf. Such species typically take readily to breeding in artificial water containers. Such casual puddles are important breeding places for some of the most serious disease vectors, such as species of Aedes that transmit dengue and yellow fever. Some with such breeding habits are disproportionately important vectors because they are well-placed to pick up pathogens from humans and pass them on. In contrast, no matter how voracious, mosquitoes that breed and feed mainly in remote wetlands and salt marshes may well remain uninfected, and if they do happen to become infected with a relevant pathogen, might seldom encounter humans to infect, in turn.
The first three stages—egg, larva, and pupa—are largely aquatic. These stages typically last 5 to 14 days, depending on the species and the ambient temperature, but there are important exceptions. Mosquitoes living in regions where some seasons are freezing or waterless spend part of the year in diapause; they delay their development, typically for months, and carry on with life only when there is enough water or warmth for their needs. For instance, Wyeomyia larvae typically get frozen into solid lumps of ice during winter and only complete their development in spring. The eggs of some species of Aedes remain unharmed in diapause if they dry out, and hatch later when they are covered by water.
Eggs hatch to become larvae, which grow until they are able to change into pupae. The adult mosquito emerges from the mature pupa as it floats at the water surface. Bloodsucking mosquitoes, depending on species, sex, and weather conditions, have potential adult lifespans ranging from as short as a week to as long as several months.
Some species can overwinter as adults in diapause.
Eggs and oviposition
Mosquito habits of oviposition, the ways in which they lay their eggs, vary considerably between species, and the morphologies of the eggs vary accordingly. The simplest procedure is that followed by many species of Anopheles; like many other gracile species of aquatic insects, females just fly over the water, bobbing up and down to the water surface and dropping eggs more or less singly. The bobbing behavior occurs among some other aquatic insects as well, for example mayflies and dragonflies; it is sometimes called "dapping". The eggs of Anopheles species are roughly cigar-shaped and have floats down their sides. Females of many common species can lay 100–200 eggs during the course of the adult phase of their lifecycles. Even with high egg and intergenerational mortality, over a period of several weeks, a single successful breeding pair can create a population of thousands.
Some other species, for example members of the genus Mansonia, lay their eggs in arrays, attached usually to the under-surfaces of waterlily pads. Their close relatives, the genus Coquillettidia, lay their eggs similarly, but not attached to plants. Instead, the eggs form layers called "rafts" that float on the water. This is a common mode of oviposition, and most species of Culex are known for the habit, which also occurs in some other genera, such as Culiseta and Uranotaenia. Anopheles eggs may on occasion cluster together on the water, too, but the clusters do not generally look much like compactly glued rafts of eggs.
In species that lay their eggs in rafts, rafts do not form adventitiously; the female Culex settles carefully on still water with its hind legs crossed, and as it lays the eggs one by one, it twitches to arrange them into a head-down array that sticks together to form the raft.
Aedes females generally drop their eggs singly, much as Anopheles do, but not as a rule into water. Instead, they lay their eggs on damp mud or other surfaces near the water's edge. Such an oviposition site commonly is the wall of a cavity such as a hollow stump or a container such as a bucket or a discarded vehicle tire. The eggs generally do not hatch until they are flooded, and they may have to withstand considerable desiccation before that happens. They are not resistant to desiccation straight after oviposition, but must develop to a suitable degree first. Once they have achieved that, however, they can enter diapause for several months if they dry out. Clutches of eggs of the majority of mosquito species hatch as soon as possible, and all the eggs in the clutch hatch at much the same time. In contrast, a batch of Aedes eggs in diapause tends to hatch irregularly over an extended period of time. This makes it much more difficult to control such species than those mosquitoes whose larvae can be killed all together as they hatch. Some Anopheles species do also behave in such a manner, though not to the same degree of sophistication.
Larva
The mosquito larva has a well-developed head with mouth brushes used for feeding, a large thorax with no legs, and a segmented abdomen.
Larvae breathe through spiracles located on their eighth abdominal segments, or through a siphon, so must come to the surface frequently. The larvae spend most of their time feeding on algae, bacteria, and other microbes in the surface microlayer.
They dive below the surface only when disturbed. Larvae swim either through propulsion with their mouth brushes, or by jerky movements of their entire bodies, giving them the common name of "wigglers" or "wrigglers".
Larvae develop through four stages, or instars, after which they metamorphose into pupae. At the end of each instar, the larvae molt, shedding their skins to allow for further growth.
Pupa
As seen in its lateral aspect, the mosquito pupa is comma-shaped. The head and thorax are merged into a cephalothorax, with the abdomen curving around underneath. The pupa can swim actively by flipping its abdomen, and it is commonly called a "tumbler" because of its swimming action. As with the larva, the pupa of most species must come to the surface frequently to breathe, which they do through a pair of respiratory trumpets on their cephalothoraxes. However, pupae do not feed during this stage; typically they pass their time hanging from the surface of the water by their respiratory trumpets. If alarmed, say by a passing shadow, they nimbly swim downwards by flipping their abdomens in much the same way as the larvae do. If undisturbed, they soon float up again.
After a few days or longer, depending on the temperature and other circumstances, the pupa rises to the water surface, the dorsal surface of its cephalothorax splits, and the adult mosquito emerges. The pupa is less active than the larva because it does not feed, whereas the larva feeds constantly.
Adult
The period of development from egg to adult varies among species and is strongly influenced by ambient temperature. Some species of mosquitoes can develop from egg to adult in as few as five days, but a more typical period of development in tropical conditions would be some 40 days or more for most species. The variation of the body size in adult mosquitoes depends on the density of the larval population and food supply within the breeding water.
Adult mosquitoes usually mate within a few days after emerging from the pupal stage. In most species, the males form large swarms, usually around dusk, and the females fly into the swarms to mate.
Males typically live for about 5–7 days, feeding on nectar and other sources of sugar. After obtaining a full blood meal, the female will rest for a few days while the blood is digested and eggs are developed. This process depends on the temperature, but usually takes two to three days in tropical conditions. Once the eggs are fully developed, the female lays them and resumes host-seeking.
The cycle repeats itself until the female dies. While females can live longer than a month in captivity, most do not live longer than one to two weeks in nature. Their lifespans depend on temperature, humidity, and their ability to successfully obtain a blood meal while avoiding host defenses and predators.
The length of the adult is typically between 3mm and 6mm. The smallest known mosquitoes are around 2 mm (0.1 in), and the largest around 19 mm (0.7 in). Mosquitoes typically weigh around 5 mg. All mosquitoes have slender bodies with three segments: a head, a thorax and an abdomen.
The head is specialized for receiving sensory information and for feeding. It has eyes and a pair of long, many-segmented antennae. The antennae are important for detecting host odors, as well as odors of breeding sites where females lay eggs. In all mosquito species, the antennae of the males in comparison to the females are noticeably bushier and contain auditory receptors to detect the characteristic whine of the females.
The compound eyes are distinctly separated from one another. Their larvae only possess a pit-eye ocellus. The compound eyes of adults develop in a separate region of the head. New ommatidia are added in semicircular rows at the rear of the eye. During the first phase of growth, this leads to individual ommatidia being square, but later in development they become hexagonal. The hexagonal pattern will only become visible when the carapace of the stage with square eyes is molted.
The head also has an elongated, forward-projecting, stinger-like proboscis used for feeding, and two sensory palps. The maxillary palps of the males are longer than their proboscises, whereas the females’ maxillary palps are much shorter. In typical bloodsucking species, the female has an elongated proboscis.
The thorax is specialized for locomotion. Three pairs of legs and a pair of wings are attached to the thorax. The insect wing is an outgrowth of the exoskeleton. The Anopheles mosquito can fly for up to four hours continuously at 1 to 2 km/h (0.6–1 mph), traveling up to 12 km (7.5 mi) in a night. Males beat their wings between 450 and 600 times per second.
The abdomen is specialized for food digestion and egg development; the abdomen of a mosquito can hold three times its own weight in blood. This segment expands considerably when a female takes a blood meal. The blood is digested over time, serving as a source of protein for the production of eggs, which gradually fill the abdomen.
Feeding by adults
Typically, both male and female mosquitoes feed on nectar and plant juices, but in many species the mouthparts of the females are adapted for piercing the skin of animal hosts and sucking their blood as ectoparasites. In many species, the female needs to obtain nutrients from a blood meal before it can produce eggs, whereas in many other species, it can produce more eggs after a blood meal. A mosquito has a variety of ways of finding its prey, including chemical, visual, and heat sensors. Both plant materials and blood are useful sources of energy in the form of sugars, and blood also supplies more concentrated nutrients, such as lipids, but the most important function of blood meals is to obtain proteins as materials for egg production.
The feeding preferences of mosquitoes include those with type O blood, heavy breathers, those with a lot of skin bacteria, people with a lot of body heat, and the pregnant. Individuals' attractiveness to mosquitoes also has a heritable, genetically-controlled component.
When a female reproduces without such parasitic meals, it is said to practice autogenous reproduction, as in Toxorhynchites; otherwise, the reproduction may be termed anautogenous, as occurs in mosquito species that serve as disease vectors, particularly Anopheles and some of the most important disease vectors in the genus Aedes. In contrast, some mosquitoes, for example, many Culex, are partially anautogenous: they do not need a blood meal for their first cycle of egg production, which they produce autogenously; however, subsequent clutches of eggs are produced anautogenously, at which point their disease vectoring activity becomes operative.
With regard to host location, female mosquitoes hunt their blood host by detecting organic substances such as carbon dioxide (CO2) and 1-octen-3-ol produced from the host, and through optical recognition. Mosquitoes prefer some people over others. The preferred victim's sweat simply smells better than others' because of the proportions of the carbon dioxide, octenol and other compounds that make up body odor. The most powerful semiochemical that triggers the keen sense of smell of Culex quinquefasciatus is nonanal. Another compound identified in human blood that attracts mosquitoes is sulcatone or 6-methyl-5-hepten-2-one, especially for Aedes aegypti mosquitoes with the odor receptor gene Or4. A large part of the mosquito’s sense of smell, or olfactory system, is devoted to sniffing out blood sources. Of 72 types of odor receptors on its antennae, at least 27 are tuned to detect chemicals found in perspiration. In Aedes, the search for a host takes place in two phases. First, the mosquito exhibits a nonspecific searching behavior until the perception of host stimulants, then it follows a targeted approach.
Most mosquito species are crepuscular (dawn or dusk) feeders. During the heat of the day, most mosquitoes rest in a cool place and wait for the evenings, although they may still bite if disturbed. Some species, such as the Asian tiger mosquito, are known to fly and feed during daytime.
Prior to and during blood feeding, blood-sucking mosquitoes inject saliva into the bodies of their source(s) of blood. This saliva serves as an anticoagulant; without it one might expect the female mosquito's proboscis to become clogged with blood clots. The saliva also is the main route by which mosquito physiology offers passenger pathogens access to the hosts' interior. The salivary glands are a major target to most pathogens, whence they find their way into the host via the stream of saliva.
The bump left on the victim's skin after a mosquito bites is called a wheal, which is caused by histamines trying to fight off the protein left by the attacking insect.
Mosquitoes of the genus Toxorhynchites never drink blood. This genus includes the largest extant mosquitoes, the larvae of which prey on the larvae of other mosquitoes. These mosquito eaters have been used in the past as mosquito control agents, with varying success.
Hosts of blood-feeding mosquito species
Many, if not all, blood-sucking species of mosquitoes are fairly selective feeders that specialise in particular host species, though they often relax their selectivity when they experience severe competition for food, defensive activity on the part of the hosts, or starvation. Some species feed selectively on monkeys, while others prefer particular kinds of birds, but they become less selective as conditions become more difficult. For example, Culiseta melanura sucks the blood of passerine birds for preference and such birds are typically the main reservoir of the Eastern equine encephalitis virus in North America. Early in the season while mosquito numbers are low, they concentrate on passerine hosts, but as mosquito numbers rise and the birds are forced to defend themselves more vigorously, the mosquitoes become less selective in attacking their avian hosts. Soon the mosquitoes begin attacking mammals more readily, thereby becoming the major vector of the virus, and causing epidemics of the disease, most conspicuously in humans and horses.
Even more dramatically, in most of its range in North America, the main vector for the Western equine encephalitis virus is Culex tarsalis, because it is known to feed variously on mammals, birds, reptiles, and amphibians. Even fish may be attacked by some mosquito species if they expose themselves above water level, as mudskippers do.
It has long been known that some species of blood-sucking flies, such as many of the Ceratopogonidae, will attack large, live insects and suck their haemolymph and that others, such as the so-called "jackal flies" (Milichiidae), will attack the recently dead prey of say, crab spiders (Thomisidae), but in the late 1960s it was reported that some species of anautogenous mosquitoes would feed on the haemolymph of caterpillars. Other observations include mosquitoes feeding on cicadas, and mantids. More recently it has been shown that malaria-transmitting mosquitoes will actively seek out some species of caterpillars and feed on their haemolymph, and do so to the caterpillar's apparent physical detriment.
Mouthparts
Mosquito mouthparts are very specialized, particularly those of the females, which in most species are adapted to piercing skin and then sucking blood. Apart from bloodsucking, the females generally also drink assorted fluids rich in dissolved sugar, such as nectar and honeydew, to obtain the energy they need. For this, their blood-sucking mouthparts are perfectly adequate. In contrast, male mosquitoes are not bloodsuckers; they only drink sugary fluids. Accordingly, their mouthparts do not require the same degree of specialization as those of females.
Externally, the most obvious feeding structure of the mosquito is the proboscis. More specifically, the visible part of the proboscis is the labium, which forms the sheath enclosing the rest of the mouthparts. When the mosquito first lands on a potential host, its mouthparts will be enclosed entirely in this sheath, and it will touch the tip of the labium to the skin in various places. Sometimes, it will begin to bite almost straight away, while other times, it will prod around, apparently looking for a suitable place. Occasionally, it will wander for a considerable time, and eventually fly away without biting. Presumably, this probing is a search for a place with easily accessible blood vessels, but the exact mechanism is not known. It is known that there are two taste receptors at the tip of the labium which may well play a role.
The female mosquito does not insert its labium into the skin; it bends back into a bow when the mosquito begins to bite. The tip of the labium remains in contact with the skin of the victim, acting as a guide for the other mouthparts. In total, there are six mouthparts besides the labium: two mandibles, two maxillae, the hypopharynx, and the labrum.
The mandibles and the maxillae are used for piercing the skin. The mandibles are pointed, while the maxillae end in flat, toothed "blades". To force these into the skin, the mosquito moves its head backwards and forwards. On one movement, the maxillae are moved as far forward as possible. On the opposite movement, the mandibles are pushed deeper into the skin by levering against the maxillae. The maxillae do not slip back because the toothed blades grip the skin.
The hypopharynx and the labrum are both hollow. Saliva with anticoagulant is pumped down the hypopharynx to prevent clotting, and blood is drawn up the labrum.
To understand the mosquito mouthparts, it is helpful to draw a comparison with an insect that chews food, such as a dragonfly. A dragonfly has two mandibles, which are used for chewing, and two maxillae, which are used to hold the food in place as it is chewed. The labium forms the floor of the dragonfly's mouth, the labrum forms the top, while the hypopharynx is inside the mouth and is used in swallowing. Conceptually, then, the mosquito's proboscis is an adaptation of the mouthparts that occur in other insects. The labium still lies beneath the other mouthparts, but also enfolds them, and it has been extended into a proboscis. The maxillae still "grip" the "food" while the mandibles "bite" it. The top of the mouth, the labrum, has developed into a channeled blade the length of the proboscis, with a cross-section like an inverted "U". Finally, the hypopharynx has extended into a tube that can deliver saliva at the end of the proboscis. Its upper surface is somewhat flattened so, when pressed against it, the labrum forms a closed tube for conveying blood from the victim.
Saliva
For the mosquito to obtain a blood meal, it must circumvent the vertebrate's physiological responses. The mosquito, as with all blood-feeding arthropods, has mechanisms to effectively block the hemostasis system with their saliva, which contains a mixture of secreted proteins. Mosquito saliva negatively affects vascular constriction, blood clotting, platelet aggregation, angiogenesis and immunity, and creates inflammation. Universally, hematophagous arthropod saliva contains at least one anti-clotting, one anti-platelet, and one vasodilatory substance. Mosquito saliva also contains enzymes that aid in sugar feeding and antimicrobial agents to control bacterial growth in the sugar meal. The composition of mosquito saliva is relatively simple, as it usually contains fewer than 20 dominant proteins. Despite the great strides in knowledge of these molecules and their role in blood feeding achieved recently, scientists still cannot ascribe functions to more than half of the molecules found in arthropod saliva. One promising application is the development of anti-clotting drugs, such as clotting inhibitors and capillary dilators, that could be useful for cardiovascular disease.
It is now well recognized that feeding ticks, sandflies, and, more recently, mosquitoes, have an ability to modulate the immune response of the animals (hosts) on which they feed. The presence of this activity in vector saliva is a reflection of the inherent overlapping and interconnected nature of the host hemostatic and inflammatory/immunological responses and the intrinsic need to prevent these host defenses from disrupting successful feeding. The mechanism for mosquito saliva-induced alteration of the host immune response is unclear, but the data have become increasingly convincing that such an effect occurs. Early work described a factor in saliva that directly suppresses TNF-α release, but not antigen-induced histamine secretion, from activated mast cells. Experiments by Cross et al. (1994) demonstrated that the inclusion of Ae. aegypti mosquito saliva into naïve cultures led to a suppression of interleukin (IL)-2 and IFN-γ production, while the cytokines IL-4 and IL-5 are unaffected by mosquito saliva. Cellular proliferation in response to IL-2 is clearly reduced by prior treatment of cells with mosquito salivary gland extract. Correspondingly, activated splenocytes isolated from mice fed upon by either Ae. aegypti or Cx. pipiens mosquitoes produce markedly higher levels of IL-4 and IL-10 concurrent with suppressed IFN-γ production. Unexpectedly, this shift in cytokine expression is observed in splenocytes up to 10 days after mosquito exposure, suggesting natural feeding of mosquitoes can have a profound, enduring, and systemic effect on the immune response.
T cell populations are decidedly susceptible to the suppressive effect of mosquito saliva, showing increased mortality and decreased division rates. Parallel work by Wasserman et al. (2004) demonstrated that T and B cell proliferation was inhibited in a dose dependent manner with concentrations as low as 1/7 of the saliva in a single mosquito. Depinay et al. (2005) observed a suppression of antibody-specific T cell responses mediated by mosquito saliva and dependent on mast cells and IL-10 expression.
A 2006 study suggests mosquito saliva can also decrease expression of interferon−α/β during early mosquito-borne virus infection. The contribution of type I interferons (IFN) in recovery from infection with viruses has been demonstrated in vivo by the therapeutic and prophylactic effects of administration of IFN inducers or IFN itself, and different research suggests mosquito saliva exacerbates West Nile virus infection, as well as other mosquito-transmitted viruses.
Egg development and blood digestion
Female mosquitoes use two very different food sources. They need sugar for energy, which is taken from sources such as nectar, and they need blood as a source of protein for egg development. Because biting is risky and hosts may be difficult to find, mosquitoes take as much blood as possible when they have the opportunity. This, however, creates another problem. Digesting that volume of blood takes a while, and the mosquito will require energy from sugar in the meantime.
To avoid this problem, mosquitoes have a digestive system which can store both food types, and give access to both as they are needed. When the mosquito drinks a sugar solution, it is directed to a crop. The crop can release sugar into the stomach as it is required. At the same time, the stomach never becomes full of sugar solution, which would prevent the mosquito taking a blood meal if it had the chance.
Blood is directed straight into the mosquito's stomach. In species that feed on mammalian or avian blood, hosts whose blood pressure is high, the mosquito feeds selectively from active blood vessels, where the pressure assists in filling the gut rapidly. If, instead of slapping a feeding mosquito, one stretches one's skin so that it grips the proboscis and the mosquito cannot withdraw it, the pressure will distend the gut until it breaks and the mosquito dies. In the unmolested mosquito, however, the mosquito will withdraw, and as the gut fills up, the stomach lining secretes a peritrophic membrane that surrounds the blood. This membrane keeps the blood separate from anything else in the stomach. However, like certain other insects that survive on dilute, purely liquid diets, notably many of the Hemiptera, many adult mosquitoes must excrete unwanted aqueous fractions even as they feed. (See the photograph of a feeding Anopheles stephensi: Note that the excreted droplet patently is not whole blood, being far more dilute). As long as they are not disturbed, this permits mosquitoes to continue feeding until they have accumulated a full meal of nutrient solids. As a result, a mosquito replete with blood can continue to absorb sugar, even as the blood meal is slowly digested over a period of several days. Once blood is in the stomach, the midgut of the female synthesizes proteolytic enzymes that hydrolyze the blood proteins into free amino acids. These are used as building blocks for the synthesis of egg yolk proteins.
In the mosquito Anopheles stephensi Liston, trypsin activity is restricted entirely to the posterior midgut lumen. No trypsin activity occurs before the blood meal, but activity increases continuously up to 30 hours after feeding, and subsequently returns to baseline levels by 60 hours. Aminopeptidase is active in the anterior and posterior midgut regions before and after feeding. In the whole midgut, activity rises from a baseline of approximately three enzyme units (EU) per midgut to a maximum of 12 EU at 30 hours after the blood meal, subsequently falling to baseline levels by 60 hours. A similar cycle of activity occurs in the posterior midgut and posterior midgut lumen, whereas aminopeptidase in the posterior midgut epithelium decreases in activity during digestion. Aminopeptidase in the anterior midgut is maintained at a constant, low level, showing no significant variation with time after feeding. Alpha-glucosidase is active in anterior and posterior midguts before and at all times after feeding. In whole midgut homogenates, alpha-glucosidase activity increases slowly up to 18 hours after the blood meal, then rises rapidly to a maximum at 30 hours after the blood meal, whereas the subsequent decline in activity is less predictable. All posterior midgut activity is restricted to the posterior midgut lumen. Depending on the time after feeding, greater than 25% of the total midgut activity of alpha-glucosidase is located in the anterior midgut. After blood meal ingestion, proteases are active only in the posterior midgut. Trypsin is the major primary hydrolytic protease and is secreted into the posterior midgut lumen without activation in the posterior midgut epithelium. Aminoptidase activity is also luminal in the posterior midgut, but cellular aminopeptidases are required for peptide processing in both anterior and posterior midguts. Alpha-glucosidase activity is elevated in the posterior midgut after feeding in response to the blood meal, whereas activity in the anterior midgut is consistent with a nectar-processing role for this midgut region.
Distribution
In the sense of the entire family Culicidae, mosquitoes are cosmopolitan; in every land region except for Antarctica and a few islands, mainly in polar or subpolar climates, at least some species of mosquito will be present. Iceland is such an island, being essentially free of mosquitoes. In warm and humid tropical regions, some mosquito species are active for the entire year, but in temperate and cold regions they hibernate or enter diapause. Arctic or subarctic mosquitoes, like some other arctic midges in families such as Simuliidae and Ceratopogonidae may be active for only a few weeks annually as melt-water pools form on the permafrost. During that time, though, they emerge in huge numbers in some regions and may take up to 300 ml of blood per day from each animal in a caribou herd.
The absence of mosquitoes from Iceland and similar regions is probably because of quirks of their climate, which differs in some respects from mainland regions. At the start of the uninterrupted continental winter of Greenland and the northern regions of Eurasia and America, the pupa enters diapause under the ice that covers sufficiently deep water. The imago ecloses only after the ice breaks in late spring. In Iceland however, the weather is less predictable. In mid-winter it frequently warms up suddenly, causing the ice to break, but then to freeze again after a few days. By that time the mosquitoes will have emerged from their pupae, but the new freeze sets in before they can complete their life cycle. Any anautogenous adult mosquito would need a host to supply a blood meal before it could lay viable eggs; it would need time to mate, mature the eggs and oviposit in suitable wetlands. These requirements would not be realistic in Iceland and in fact the absence of mosquitoes from such subpolar islands is in line with the islands' low biodiversity; Iceland has fewer than 1500 described species of insects, many of them probably accidentally introduced by human agency. In Iceland most ectoparasitic insects live in sheltered conditions or actually on mammals; examples include lice, fleas and bedbugs, in whose living conditions freezing is no concern, and most of which were introduced inadvertently by humans.
Some other aquatic Diptera, such as Simuliidae, do survive in Iceland, but their habits and adaptations differ from those of mosquitoes; Simuliidae for example, though they, like mosquitoes, are bloodsuckers, generally inhabit stones under running water that does not readily freeze and which is totally unsuited to mosquitoes; mosquitoes are generally not adapted to running water.
Eggs of species of mosquitoes from the temperate zones are more tolerant of cold than the eggs of species indigenous to warmer regions. Many even tolerate subzero temperatures. In addition, adults of some species can survive the winter by taking shelter in suitable microhabitats such as buildings or hollow trees.
Means of dispersal
Worldwide introduction of various mosquito species over large distances into regions where they are not indigenous has occurred through human agencies, primarily on sea routes, in which the eggs, larvae, and pupae inhabiting water-filled used tires and cut flowers are transported. However, apart from sea transport, mosquitoes have been effectively carried by personal vehicles, delivery trucks, trains, and aircraft. Man-made areas such as storm water retention basins, or storm drains also provide sprawling sanctuaries. Sufficient quarantine measures have proven difficult to implement. In addition, outdoor pool areas make a perfect place for them to grow.
Disease
Mosquitoes can act as vectors for many disease-causing viruses and parasites. Infected mosquitoes carry these organisms from person to person without exhibiting symptoms themselves. Mosquito-borne diseases include:
Potential transmission of HIV was originally a public health concern, but practical considerations and detailed studies of epidemiological patterns suggest that any transmission of the HIV virus by mosquitoes is at worst extremely unlikely.
Various species of mosquitoes are estimated to transmit various types of disease to more than 700 million people annually in Africa, South America, Central America, Mexico, Russia, and much of Asia, with millions of resultant deaths. At least two million people annually die of these diseases, and the morbidity rates are many times higher still.
Methods used to prevent the spread of disease, or to protect individuals in areas where disease is endemic, include:
Since most such diseases are carried by "elderly" female mosquitoes, some scientists have suggested focusing on these to avoid the evolution of resistance.
Indiscriminate eradication of mosquitoes is likely to have effects undesirable to humans. Entomologist Phil Lounibos of Florida Medical Entomological Laboratory (FMEL), Institute of Food and Agricultural Sciences (IFAS), University of Florida, says that eradication "is fraught with undesirable side effects", as mosquitoes are a significant food source for birds and bats and, as larvae, fish and frogs, parts of the food chain affecting many species. If mosquitoes are eradicated, they may also be replaced by other species, possibly more undesirable.
Control
Many measures have been tried for mosquito control, including the elimination of breeding places, exclusion via window screens and mosquito nets, biological control with parasites such as fungi and nematodes, or predators such as fish, copepods, dragonfly nymphs and adults, and some species of lizard and gecko. Another approach is to introduce large numbers of sterile males. Genetic methods including cytoplasmic incompatibility, chromosomal translocations, sex distortion and gene replacement have been explored. They are cheaper and not subject to vector resistance.
Repellents
Insect repellents are applied on skin and give short-term protection against mosquito bites. The chemical DEET repels some mosquitoes and other insects. Some CDC-recommended repellents are picaridin, eucalyptus oil (PMD) and IR3535. Others are indalone, dimethyl phthalate, dimethyl carbate, and ethyl hexanediol. More recently, in 2015, Researchers at New Mexico State University tested 10 commercially available products for their effectiveness at repelling mosquitoes. On the mosquito Aedes aegypti, the vector of Zika virus, only one repellent that did not contain DEET had a strong effect for the duration of the 240 minutes test: a lemon eucalyptus oil repellent. All DEET-containing mosquito repellents were active.
There are also electronic insect repellent devices which produce ultrasounds that were developed to keep away insects (and mosquitoes). However, no scientific research based on the EPA's and many universities' studies has ever provided evidence that these devices prevent a human from being bitten by a mosquito. In 2005, the British consumer magazine Holiday reported the results of its test of a range of mosquito deterrents. The magazine's editor Lorna Cowan described the four appliances that used a buzzer as "a shocking waste of money" which "should be removed from sale".
Bites and treatment
Visible, irritating bites are due to an immune response from the binding of IgG and IgE antibodies to antigens in the mosquito's saliva. Some of the sensitizing antigens are common to all mosquito species, whereas others are specific to certain species. There are both immediate hypersensitivity reactions (types I and III) and delayed hypersensitivity reactions (type IV) to mosquito bites. Both reactions result in itching, redness and swelling. Immediate reactions develop within a few minutes of the bite and last for a few hours. Delayed reactions take around a day to develop, and last for up to a week. Several anti-itch medications are commercially available, including those taken orally, such as diphenhydramine, or topically applied antihistamines and, for more severe cases, corticosteroids, such as hydrocortisone and triamcinolone.
In human culture
Ancient Greek beast fables including "The Elephant and the Mosquito" and "The Bull and the Mosquito", with the general moral that the large beast does not even notice the small one, derive ultimately from Mesopotamia.
Winsor McCay's 1912 film How a Mosquito Operates was one of the earliest works of animation, far ahead of its time in technical quality. It depicts a giant mosquito tormenting a sleeping man.
The de Havilland Mosquito was a high-speed aircraft manufactured between 1940 and 1950, and used in many roles.