
 | ||
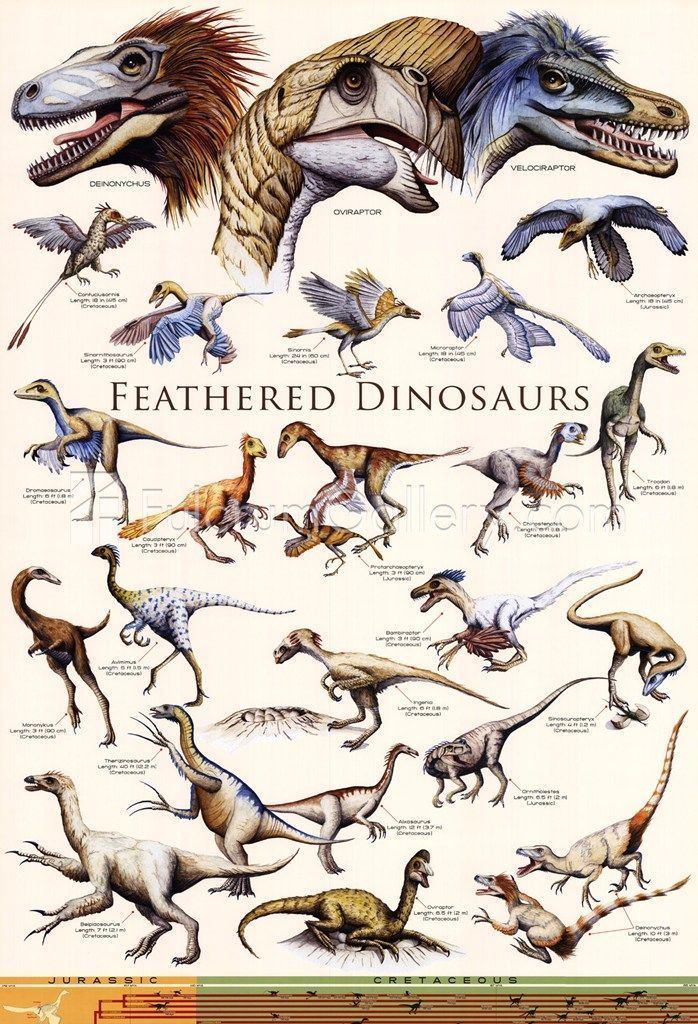
Representative species | ||
A feathered dinosaur is any species of dinosaur possessing feathers. For over 150 years, since scientific research began on dinosaurs in the early 1800s, dinosaurs were generally believed to be related to the reptile family; the word "dinosaur", coined in 1842 by paleontologist Richard Owen, comes from the Greek for "formidable lizard". This view began to shift during the so-called dinosaur renaissance in scientific research in the late 1960s, and by the mid-1990s significant evidence had emerged that dinosaurs are much more closely related to birds. In fact, birds are now believed to have descended directly from the theropod group of dinosaurs, and are thus classified as dinosaurs themselves, meaning that any modern bird can be considered a feathered dinosaur, since all modern birds possess feathers (with the exception of a few artificially selected chickens).
Contents
- Feathered dinosaurs bird ancestors
- Early
- Fossil discoveries
- List of non avian dinosaur species preserved with evidence of feathers
- Primitive feather types
- Phylogeny and the inference of feathers in other dinosaurs
- References

Among extinct dinosaurs, feathers or feather-like integument have been discovered on dozens of genera via both direct and indirect fossil evidence. The vast majority of feather discoveries have been for coelurosaurian theropods. However, integument has also been discovered on at least three ornithischians, raising the likelihood that proto-feathers were also present in earlier dinosaurs, and perhaps even a more ancestral animal, in light of the pycnofibers of pterosaurs. Crocodilians also possess beta keratin very similar to those of birds, which suggests that they evolved from a common ancestral gene.

Feathered dinosaurs bird ancestors
Early
Shortly after the 1859 publication of Charles Darwin's On the Origin of Species, British biologist Thomas Henry Huxley proposed that birds were descendants of dinosaurs. He compared the skeletal structure of Compsognathus, a small theropod dinosaur, and the 'first bird' Archaeopteryx lithographica (both of which were found in the Upper Jurassic Bavarian limestone of Solnhofen). He showed that, apart from its hands and feathers, Archaeopteryx was quite similar to Compsognathus. Thus Archaeopteryx represents a transitional fossil. In 1868 he published On the Animals which are most nearly intermediate between Birds and reptiles, making the case. The first restoration of a feathered dinosaur was Thomas Henry Huxley's depiction in 1876 of a feathered Compsognathus to accompany a lecture on the evolution of birds he delivered in New York in which he speculated that the aforementioned dinosaur might have been in possession of feathers. The leading dinosaur expert of the time, Richard Owen, disagreed, claiming Archaeopteryx as the first bird outside dinosaur lineage. For the next century, claims that birds were dinosaur descendants faded, with more popular bird-ancestry hypotheses including 'crocodylomorph' and 'thecodont' ancestors, rather than dinosaurs or other archosaurs.

In 1969, John Ostrom described Deinonychus antirrhopus, a theropod that he had discovered in Montana in 1964 and whose skeletal resemblance to birds seemed unmistakable. Ostrom became a leading proponent of the theory that birds are direct descendants of dinosaurs. Further comparisons of bird and dinosaur skeletons, as well as cladistic analysis strengthened the case for the link, particularly for a branch of theropods called maniraptors. Skeletal similarities include the neck, the pubis, the wrists (semi-lunate carpal), the 'arms' and pectoral girdle, the shoulder blade, the clavicle and the breast bone. In all, over a hundred distinct anatomical features are shared by birds and theropod dinosaurs.

Other researchers drew on these shared features and other aspects of dinosaur biology and began to suggest that at least some theropod dinosaurs were feathered. Gregory S. Paul was probably among the first paleoartists to depict non-avian maniraptoran dinosaurs with feathers and protofeathers, starting in the late 1970s.
By the 1990s, most paleontologists considered birds to be surviving dinosaurs and referred to 'non-avian dinosaurs' (all extinct), to distinguish them from birds (Avialae). Before the discovery of feathered dinosaurs, the evidence was limited to Huxley and Ostrom's comparative anatomy. Some mainstream ornithologists, including Smithsonian Institution curator Storrs L. Olson, disputed the links, specifically citing the lack of fossil evidence for feathered dinosaurs.
Fossil discoveries
After a century of hypotheses without conclusive evidence, well-preserved fossils of feathered dinosaurs were discovered during the 1990s, and more continue to be found. The fossils were preserved in a Lagerstätte — a sedimentary deposit exhibiting remarkable richness and completeness in its fossils — in Liaoning, China. The area had repeatedly been smothered in volcanic ash produced by eruptions in Inner Mongolia 124 million years ago, during the Early Cretaceous epoch. The fine-grained ash preserved the living organisms that it buried in fine detail. The area was teeming with life, with millions of leaves, angiosperms (the oldest known), insects, fish, frogs, salamanders, mammals, turtles, and lizards discovered to date.
The most important discoveries at Liaoning have been a host of feathered dinosaur fossils, with a steady stream of new finds filling in the picture of the dinosaur–bird connection and adding more to theories of the evolutionary development of feathers and flight. Turner et al. (2007) reported quill knobs from an ulna of Velociraptor mongoliensis, and these are strongly correlated with large and well-developed secondary feathers.
Behavioural evidence, in the form of an oviraptorosaur on its nest, showed another link with birds. Its forearms were folded, like those of a bird. Although no feathers were preserved, it is likely that these would have been present to insulate eggs and juveniles.
Not all of the Chinese fossil discoveries proved valid however. In 1999, a supposed fossil of an apparently feathered dinosaur named Archaeoraptor liaoningensis, found in Liaoning Province, northeastern China, turned out to be a forgery. Comparing the photograph of the specimen with another find, Chinese paleontologist Xu Xing came to the conclusion that it was composed of two portions of different fossil animals. His claim made National Geographic review their research and they too came to the same conclusion. The bottom portion of the "Archaeoraptor" composite came from a legitimate feathered dromaeosaurid now known as Microraptor, and the upper portion from a previously-known primitive bird called Yanornis.
In 2011, samples of amber were discovered to contain preserved feathers from 75 to 80 million years ago during the Cretaceous era, with evidence that they were from both dinosaurs and birds. Initial analysis suggests that some of the feathers were used for insulation, and not flight. More complex feathers were revealed to have variations in coloration similar to modern birds, while simpler protofeathers were predominantly dark. Only 11 specimens are currently known. The specimens are too rare to be broken open to study their melanosomes, but there are plans for using non-destructive high-resolution X-ray imaging.
In 2016, the discovery was announced of a feathered dinosaur tail preserved in amber that is estimated to be 99 million years old. Lida Xing, a researcher from the China University of Geosciences in Beijing, found the specimen at an amber market in Myanmar. It is the first definitive discovery of dinosaur material in amber.
List of non-avian dinosaur species preserved with evidence of feathers
A number of non-avian dinosaurs are now known to have been feathered. Direct evidence of feathers exists for the following species, listed in the order currently accepted evidence was first published. In all examples, the evidence described consists of feather impressions, except those genera inferred to have had feathers based on skeletal or chemical evidence, such as the presence of quill knobs (the anchor points for wing feathers on the forelimb) or a pygostyle (the fused vertebrae at the tail tip which often supports large feathers).
- Avimimus portentosus (inferred 1987: ulnar ridge)
- Sinosauropteryx prima (1996)
- Protarchaeopteryx robusta (1997)
- GMV 2124 (1997)
- Caudipteryx zoui (1998)
- Rahonavis ostromi (inferred 1998: quill knobs; possibly avialan)
- Shuvuuia deserti (1999)
- Beipiaosaurus inexpectus (1999)
- Sinornithosaurus millenii (1999)
- Caudipteryx dongi (2000)
- Caudipteryx sp. (2000)
- Microraptor zhaoianus (2000)
- Nomingia gobiensis (inferred 2000: pygostyle)
- Psittacosaurus sp.? (2002)
- Scansoriopteryx heilmanni (2002; possibly avialan)
- Yixianosaurus longimanus (2003)
- Dilong paradoxus (2004)
- Pedopenna daohugouensis (2005; possibly avialan)
- Jinfengopteryx elegans (2005)
- Juravenator starki (2006)
- Sinocalliopteryx gigas (2007)
- Velociraptor mongoliensis (inferred 2007: quill knobs)
- Epidexipteryx hui (2008; possibly avialan)
- Similicaudipteryx yixianensis (inferred 2008: pygostyle; confirmed 2010)
- Anchiornis huxleyi (2009; possibly avialan)
- Tianyulong confuciusi? (2009)
- Concavenator corcovatus? (inferred 2010: quill knobs?)
- Xiaotingia zhengi (2011; possibly avialan)
- Yutyrannus huali (2012)
- Sciurumimus albersdoerferi (2012)
- Ornithomimus edmontonicus (2012)
- Ningyuansaurus wangi (2012)
- Eosinopteryx brevipenna (2013; possibly avialan)
- Jianchangosaurus yixianensis (2013)
- Aurornis xui (2013; possibly avialan)
- Changyuraptor yangi (2014)
- Kulindadromeus zabaikalicus? (2014)
- Citipati osmolskae (inferred 2014: pygostyle)
- Conchoraptor gracilis (inferred 2014: pygostyle)
- Deinocheirus mirificus? (inferred 2014: pygostyle)
- Yi qi (2015)
- Zhenyuanlong suni (2015)
- Dakotaraptor steini (inferred 2015: quill knobs)
- Apatoraptor pennatus (inferred 2016: quill knobs)
Primitive feather types
Integumentary structures that gave rise to the feathers of birds are seen in the dorsal spines of reptiles and fish. A similar stage in their evolution to the complex coats of birds and mammals can be observed in living reptiles such as iguanas and Gonocephalus agamids. Feather structures are thought to have proceeded from simple hollow filaments through several stages of increasing complexity, ending with the large, deeply rooted feathers with strong pens (rachis), barbs and barbules that birds display today.
According to Prum's (1999) proposed model, at stage I, the follicle originates with a cylindrical epidermal depression around the base of the feather papilla. The first feather resulted when undifferentiated tubular follicle collar developed out of the old keratinocytes being pushed out. At stage II, the inner, basilar layer of the follicle collar differentiated into longitudinal barb ridges with unbranched keratin filaments, while the thin peripheral layer of the collar became the deciduous sheath, forming a tuft of unbranched barbs with a basal calamus. Stage III consists of two developmental novelties, IIIa and IIIb, as either could have occurred first. Stage IIIa involves helical displacement of barb ridges arising within the collar. The barb ridges on the anterior midline of the follicle fuse together, forming the rachis. The creation of a posterior barb locus follows, giving an indeterminate number of barbs. This resulted in a feather with a symmetrical, primarily branched structure with a rachis and unbranched barbs. In stage IIIb, barbules paired within the peripheral barbule plates of the barb ridges, create branched barbs with rami and barbules. This resulting feather is one with a tuft of branched barbs without a rachis. At stage IV, differentiated distal and proximal barbules produce a closed, pennaceous vane. A closed vane develops when pennulae on the distal barbules form a hooked shape to attach to the simpler proximal barbules of the adjacent barb. Stage V developmental novelties gave rise to additional structural diversity in the closed pennaceous feather. Here, asymmetrical flight feathers, bipinnate plumulaceous feathers, filoplumes, powder down, and bristles evolved.
Some evidence suggests that the original function of simple feathers was insulation. In particular, preserved patches of skin in large, derived, tyrannosauroids show scutes, while those in smaller, more primitive, forms show feathers. This may indicate that the larger forms had complex skins, with both scutes and filaments, or that tyrannosauroids may be like rhinos and elephants, having filaments at birth and then losing them as they developed to maturity. An adult Tyrannosaurus rex weighed about as much as an African elephant. If large tyrannosauroids were endotherms, they would have needed to radiate heat efficiently. However, due to the different structural properties of feathers compared to fur, as well as a larger surface area per cubic square meter, it is extremely unlikely even the largest theropods would suffer overheating issues from an extensive feather coat.
There is an increasing body of evidence that supports the display hypothesis, which states that early feathers were colored and increased reproductive success. Coloration could have provided the original adaptation of feathers, implying that all later functions of feathers, such as thermoregulation and flight, were co-opted. This hypothesis has been supported by the discovery of pigmented feathers in multiple species. Supporting the display hypothesis is the fact that fossil feathers have been observed in a ground-dwelling herbivorous dinosaur clade, making it unlikely that feathers functioned as predatory tools or as a means of flight. Additionally, some specimens have iridescent feathers. Pigmented and iridescent feathers may have provided greater attractiveness to mates, providing enhanced reproductive success when compared to non-colored feathers. Current research shows that it is plausible that theropods would have had the visual acuity necessary to see the displays. In a study by Stevens (2006), the binocular field of view for Velociraptor has been estimated to be 55 to 60 degrees, which is about that of modern owls. Visual acuity for Tyrannosaurus has been predicted to be anywhere from about that of humans to 13 times that of humans. However, as both Velociraptor and Tyrannosaurus have a rather extended evolutionary relationship with the more basal theropods, it is unclear how much of this visual acuity data can be extrapolated.
The fact that precursors of feathers appeared and then were co-opted for insulation is already present in Gould and Vrba, 1982. The reason why such precursors appeared could be explained by a theory which is based on metabolic issue. Feathers are made of protein and contain substantial amounts of certain amino acids, especially cysteine. The protein complex at the base of the composition of the feather is keratin, which has disulfide bonds between amino acids that confer unique properties of stability and elasticity. The metabolism of amino acids containing sulfur proved to be toxic to the organism. If the sulfur amino acids are not catabolized at the final products of urea or uric acid but used for the synthesis of keratin instead, the release of hydrogen sulfide is extremely reduced or avoided. For an organism whose metabolism works at high internal temperatures of 40 °C or greater can be extremely important to prevent the excess production of hydrogen sulfide. This hypothesis could be consistent with the need for high metabolic rate of theropod dinosaurs.
It is not known with certainty at what point in archosaur phylogeny the earliest simple "protofeathers" arose, or whether they arose once or independently multiple times. Filamentous structures are clearly present in pterosaurs, and long, hollow quills have been reported in specimens of the ornithischian dinosaurs Psittacosaurus and Tianyulong. In 2009, Xu et al. noted that the hollow, unbranched, stiff integumentary structures found on a specimen of Beipiaosaurus were strikingly similar to the integumentary structures of Psittacosaurus and pterosaurs. They suggested that all of these structures may have been inherited from a common ancestor much earlier in the evolution of archosaurs, possibly in an ornithodire from the Middle Triassic or earlier. More recently, findings in Russia of the basal neornithischian Kulindadromeus report that although the lower leg and tail seemed to be scaled, "varied integumentary structures were found directly associated with skeletal elements, supporting the hypothesis that simple filamentous feathers, as well as compound feather-like structures comparable to those in theropods, were widespread amongst the whole dinosaur clade."
Display feathers are also known from dinosaurs that are very primitive members of the bird lineage, or Avialae. The most primitive example is Epidexipteryx, which had a short tail with extremely long, ribbon-like feathers. Oddly enough, the fossil does not preserve wing feathers, suggesting that Epidexipteryx was either secondarily flightless, or that display feathers evolved before flight feathers in the bird lineage. Plumaceous feathers are found in nearly all lineages of Theropoda common in the northern hemisphere, and pennaceous feathers are attested as far down the tree as the Ornithomimosauria. The fact that only adult Ornithomimus had wing-like structures suggests that pennaceous feathers evolved for mating displays.
Phylogeny and the inference of feathers in other dinosaurs
Fossil feather impressions are extremely rare and they require exceptional preservation conditions to form. Therefore, only a few feathered dinosaur genera have been identified. All fossil feather specimens have been found to show certain similarities. Due to these similarities and through developmental research, almost all scientists agree that feathers could only have evolved once in dinosaurs. Feathers would then have been passed down to all later, more derived species, unless some lineages lost feathers secondarily. If a dinosaur falls at a point on an evolutionary tree within the known feather-bearing lineages, then its ancestors had feathers, and it is quite possible that it did as well. This technique, called phylogenetic bracketing, can also be used to infer the type of feathers a species may have had, since the developmental history of feathers is now reasonably well-known. All feathered species had filamentaceous or plumaceous (downy) feathers, with pennaceous feathers found among the more bird-like groups. The following cladogram is adapted from Godefroit et al., 2013.
Grey denotes a clade that is not known to contain any feathered specimen at the time of writing (although this does not imply that members of the clade lacked feathers).
|label1=Neotheropoda |1=