
Scientific name Tyrannosaurus Higher classification Tyrannosaurini | Subfamily †Tyrannosaurinae Extinction status Extinct | |
 | ||
Speed Tyrannosaurus rex: 40 km/h Similar dinosaur, Triceratops, Spinosaurus, Velociraptor, Stegosaurus | ||
tyrannosaurus rex dinosaurs songs about the t rex by storybots
Tyrannosaurus (/TᵻˌræNəˈSɔːrəs/ or /taɪˌræNəˈsɔːrəs/, meaning "tyrant lizard", from the Ancient Greek tyrannos (τύραννος), "tyrant", and sauros (σαῦρος), "lizard") is a genus of coelurosaurian theropod dinosaur. The species Tyrannosaurus rex (rex meaning "king" in Latin), is one of the most well-represented of the large theropods. Tyrannosaurus lived throughout what is now western North America, on what was then an island continent known as Laramidia. Tyrannosaurus had a much wider range than other tyrannosaurids. Fossils are found in a variety of rock formations dating to the Maastrichtian age of the upper Cretaceous Period, 68 to 66 million years ago. It was the last known member of the tyrannosaurids, and among the last non-avian dinosaurs to exist before the Cretaceous–Paleogene extinction event.
Contents
- tyrannosaurus rex dinosaurs songs about the t rex by storybots
- Description
- Skin and feathers
- Classification
- Life history
- Sexual dimorphism
- Posture
- Arms
- Soft tissue
- Thermoregulation
- Footprints
- Locomotion
- Brain and senses
- Feeding strategies
- Cannibalism
- Pack behavior
- Pathology
- Paleoecology
- History of research
- Earliest finds
- Manospondylus
- Notable specimens
- In popular culture
- References

Like other tyrannosaurids, Tyrannosaurus was a bipedal carnivore with a massive skull balanced by a long, heavy tail. Relative to its large and powerful hind limbs, Tyrannosaurus fore limbs were short but unusually powerful for their size and had two clawed digits. The most complete specimen measures up to 12.3 m (40 ft) in length, up to 3.66 meters (12 ft) tall at the hips, and according to most modern estimates 8.4 metric tons (9.3 short tons) to 18.5 metric tons (20.4 short tons) in weight. Although other theropods rivaled or exceeded Tyrannosaurus rex in size, it is still among the largest known land predators and is estimated to have exerted the largest bite force among all terrestrial animals. By far the largest carnivore in its environment, Tyrannosaurus rex was most likely an apex predator, preying upon hadrosaurs, armoured herbivores like ceratopsians and ankylosaurs, and possibly sauropods. Some experts, however, have suggested the dinosaur was primarily a scavenger. The question of whether Tyrannosaurus was an apex predator or a pure scavenger was among the longest ongoing debates in paleontology. It is accepted now that Tyrannosaurus rex acted as a predator, and opportunistically scavenged as modern mammalian and avian predators do.

More than 50 specimens of Tyrannosaurus rex have been identified, some of which are nearly complete skeletons. Soft tissue and proteins have been reported in at least one of these specimens. The abundance of fossil material has allowed significant research into many aspects of its biology, including its life history and biomechanics. The feeding habits, physiology and potential speed of Tyrannosaurus rex are a few subjects of debate. Its taxonomy is also controversial, as some scientists consider Tarbosaurus bataar from Asia to be a second Tyrannosaurus species while others maintain Tarbosaurus is a separate genus. Several other genera of North American tyrannosaurids have also been synonymized with Tyrannosaurus.
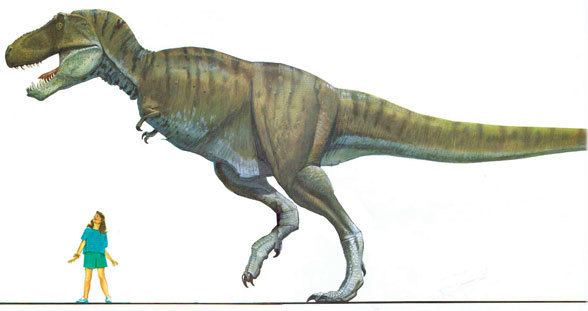
Description

Tyrannosaurus rex was one of the largest land carnivores of all time; the largest complete specimen, located at the Field Museum of Natural History under the name FMNH PR2081 and nicknamed Sue, measured 12.3 meters (40 ft) long, and was 3.66 meters (12 ft) tall at the hips, and according to the most recent studies estimated to have weighed between 8.4 metric tons (9.3 short tons) to 18.5 metric tons (20.4 short tons) when alive. However, not every adult Tyrannosaurus specimen recovered is as big. Historically average adult mass estimates have varied widely over the years, from as low as 4.5 metric tons (5.0 short tons), to more than 7.2 metric tons (7.9 short tons), with most modern estimates ranging between 5.4 metric tons (6.0 short tons) and 8.0 metric tons (8.8 short tons). Hutchinson et al. (2011) found that the maximum weight of Sue, the largest complete Tyrannosaurus specimen, was between 9.5 and 18.5 metric tons (9.3–18.2 long tons; 10.5–20.4 short tons), though the authors stated that their upper and lower estimates were based on models with wide error bars and that they "consider [them] to be too skinny, too fat, or too disproportionate" and provided a mean estimate at 13.99 metric tons (15.42 short tons) for this specimen. Packard et al. (2009) tested dinosaur mass estimation procedures on elephants and concluded that those of dinosaurs are flawed and produce over-estimations; thus, the weight of Tyrannosaurus, as well as other dinosaurs, could have been much less. Other estimations have concluded that the largest known Tyrannosaurus specimens had masses approaching or exceeding 9 tonnes.

Due to the relatively small number of recovered specimens and the large population of individuals present at any given time when Tyrannosaurus was alive, there could have easily been larger specimens than those currently known including "Sue", though discovery of these largest individuals may be forever untenable due to the incomplete nature of the fossil record. Holtz has also suggested that "it is very reasonable to suspect that there were individuals that were 10, 15, or even 20 percent larger than Sue in any T. rex population."
The neck of Tyrannosaurus rex formed a natural S-shaped curve like that of other theropods, but was short and muscular to support the massive head. The forelimbs had only two clawed fingers, along with an additional small metacarpal representing the remnant of a third digit. In contrast the hind limbs were among the longest in proportion to body size of any theropod. The tail was heavy and long, sometimes containing over forty vertebrae, in order to balance the massive head and torso. To compensate for the immense bulk of the animal, many bones throughout the skeleton were hollow, reducing its weight without significant loss of strength.
The largest known Tyrannosaurus rex skulls measure up to 1.45 meters (4.8 ft) in length. Large fenestrae (openings) in the skull reduced weight and provided areas for muscle attachment, as in all carnivorous theropods. But in other respects Tyrannosaurus's skull was significantly different from those of large non-tyrannosauroid theropods. It was extremely wide at the rear but had a narrow snout, allowing unusually good binocular vision. The skull bones were massive and the nasals and some other bones were fused, preventing movement between them; but many were pneumatized (contained a "honeycomb" of tiny air spaces) which may have made the bones more flexible as well as lighter. These and other skull-strengthening features are part of the tyrannosaurid trend towards an increasingly powerful bite, which easily surpassed that of all non-tyrannosaurids. The tip of the upper jaw was U-shaped (most non-tyrannosauroid carnivores had V-shaped upper jaws), which increased the amount of tissue and bone a tyrannosaur could rip out with one bite, although it also increased the stresses on the front teeth.
The teeth of Tyrannosaurus rex displayed marked heterodonty (differences in shape). The premaxillary teeth at the front of the upper jaw were closely packed, D-shaped in cross-section, had reinforcing ridges on the rear surface, were incisiform (their tips were chisel-like blades) and curved backwards. The D-shaped cross-section, reinforcing ridges and backwards curve reduced the risk that the teeth would snap when Tyrannosaurus bit and pulled. The remaining teeth were robust, like "lethal bananas" rather than daggers, more widely spaced and also had reinforcing ridges. Those in the upper jaw were larger than those in all but the rear of the lower jaw. The largest found so far is estimated to have been 30.5 centimeters (12 in) long including the root when the animal was alive, making it the largest tooth of any carnivorous dinosaur yet found.
Skin and feathers
While there is no direct evidence for Tyrannosaurus rex having had feathers, many scientists now consider it likely that T. rex had feathers on at least parts of its body, due to their presence in related species. Mark Norell of the American Museum of Natural History summarized the balance of evidence by stating that: "we have as much evidence that T. rex was feathered, at least during some stage of its life, as we do that australopithecines like Lucy had hair."
The first evidence for feathers in tyrannosauroids came from the small species Dilong paradoxus, found in the Yixian Formation of China, and reported in 2004. As with many other theropods discovered in the Yixian, the fossil skeleton was preserved with a coat of filamentous structures which are commonly recognized as the precursors of feathers. Because all known skin impressions from larger tyrannosauroids known at the time showed evidence of scales, the researchers who studied Dilong speculated that feathers may correlate negatively with body size—that juveniles may have been feathered, then shed the feathers and expressed only scales as the animal became larger and no longer needed insulation to stay warm. However, subsequent discoveries showed that even some large tyrannosauroids had feathers covering much of their bodies, casting doubt on the hypothesis that they were a size-related feature.
While skin impressions from a Tyrannosaurus rex specimen nicknamed "Wyrex" (BHI 6230) discovered in Montana in 2002, as well as some other giant tyrannosauroid specimens, show at least small patches of mosaic scales, others, such as Yutyrannus huali (which was up to 9 meters (30 ft) long and weighed about 1,400 kilograms (3,100 lb)), preserve feathers on various sections of the body, strongly suggesting that its whole body was covered in feathers. It is possible that the extent and nature of feather covering in tyrannosauroids may have changed over time in response to body size, a warmer climate, or other factors.
Research has suggested that large theropods like Tyrannosaurus had teeth covered in lips like modern day lizards, rather than having bare teeth like crocodiles. This is based on the presence of enamel, which needs to remain hydrated, an issue not faced by aquatic species like crocodilians or toothless species like birds, which have lipless mouths and toothless beaks. In modern terrestrial animals with bare teeth, the teeth typically lack some or all of the standard amount of tooth enamel. Because Tyrannosaurus and other toothed theropods had enamel-covered teeth, they would have required some sort of lip-like covering to seal the mouth when closed.
Classification
Tyrannosaurus is the type genus of the superfamily Tyrannosauroidea, the family Tyrannosauridae, and the subfamily Tyrannosaurinae; in other words it is the standard by which paleontologists decide whether to include other species in the same group. Other members of the tyrannosaurine subfamily include the North American Daspletosaurus and the Asian Tarbosaurus, both of which have occasionally been synonymized with Tyrannosaurus. Tyrannosaurids were once commonly thought to be descendants of earlier large theropods such as megalosaurs and carnosaurs, although more recently they were reclassified with the generally smaller coelurosaurs.
In 1955, Soviet paleontologist Evgeny Maleev named a new species, Tyrannosaurus bataar, from Mongolia. By 1965, this species had been renamed Tarbosaurus bataar. Despite the renaming, many phylogenetic analyses have found Tarbosaurus bataar to be the sister taxon of Tyrannosaurus rex, and it has often been considered an Asian species of Tyrannosaurus. A recent redescription of the skull of Tarbosaurus bataar has shown that it was much narrower than that of Tyrannosaurus rex and that during a bite, the distribution of stress in the skull would have been very different, closer to that of Alioramus, another Asian tyrannosaur. A related cladistic analysis found that Alioramus, not Tyrannosaurus, was the sister taxon of Tarbosaurus, which, if true, would suggest that Tarbosaurus and Tyrannosaurus should remain separate. The discovery and description of Qianzhousaurus would later disprove this and revealed that Alioramus belonged to the clade Alioramini. The discovery of the tyrannosaurid Lythronax further indicates that Tarbosaurus and Tyrannosaurus are closely related, forming a clade with fellow Asian tyrannosaurid Zhuchengtyrannus, with Lythronax being their sister taxon. A further study from 2016 by Steve Brusatte, Thomas Carr et al., also indicates Tyrannosaurus may have been an immigrant from Asia, as well as a possible descendent of Tarbosaurus. The study further indicates the possibility that Tyrannosaurus may have driven other tyrannosaurids that were native to North America extinct through competition. Other finds in 2006 indicate giant tyrannosaurs may have been present in North America as early as 75 million years ago. Whether or not this specimen belongs to Tyrannosaurus rex, a new species of Tyrannosaurus, or a new genus entirely is still unknown.
Other tyrannosaurid fossils found in the same formations as Tyrannosaurus rex were originally classified as separate taxa, including Aublysodon and Albertosaurus megagracilis, the latter being named Dinotyrannus megagracilis in 1995. However, these fossils are now universally considered to belong to juvenile Tyrannosaurus rex. A small but nearly complete skull from Montana, 60 centimeters (2.0 ft) long, may be an exception. This skull was originally classified as a species of Gorgosaurus (G. lancensis) by Charles W. Gilmore in 1946, but was later referred to a new genus, Nanotyrannus. Opinions remain divided on the validity of N. lancensis. Many paleontologists consider the skull to belong to a juvenile Tyrannosaurus rex. There are minor differences between the two species, including the higher number of teeth in N. lancensis, which lead some scientists to recommend keeping the two genera separate until further research or discoveries clarify the situation.
Below is the cladogram of Tyrannosauridae based on the phylogenetic analysis conducted by Loewen et al. in 2013.
Life history
The identification of several specimens as juvenile Tyrannosaurus rex has allowed scientists to document ontogenetic changes in the species, estimate the lifespan, and determine how quickly the animals would have grown. The smallest known individual (LACM 28471, the "Jordan theropod") is estimated to have weighed only 30 kg (66 lb), while the largest, such as FMNH PR2081 (Sue) most likely weighed about 5,650 kg (12,460 lb). Histologic analysis of Tyrannosaurus rex bones showed LACM 28471 had aged only 2 years when it died, while Sue was 28 years old, an age which may have been close to the maximum for the species.
Histology has also allowed the age of other specimens to be determined. Growth curves can be developed when the ages of different specimens are plotted on a graph along with their mass. A Tyrannosaurus rex growth curve is S-shaped, with juveniles remaining under 1,800 kg (4,000 lb) until approximately 14 years of age, when body size began to increase dramatically. During this rapid growth phase, a young Tyrannosaurus rex would gain an average of 600 kg (1,300 lb) a year for the next four years. At 18 years of age, the curve plateaus again, indicating that growth slowed dramatically. For example, only 600 kg (1,300 lb) separated the 28-year-old Sue from a 22-year-old Canadian specimen (RTMP 81.12.1). A 2004 histological study performed by different workers corroborates these results, finding that rapid growth began to slow at around 16 years of age. Another study corroborated the latter study's results but found the growth rate to be much faster, finding it to be around 1800 kilograms (4000 lbs). Although these results were much higher than previous estimations, the authors noted that these results significantly lowered the great difference between its actual growth rate and the one which would be expected of an animal of its size. The sudden change in growth rate at the end of the growth spurt may indicate physical maturity, a hypothesis which is supported by the discovery of medullary tissue in the femur of a 16 to 20-year-old Tyrannosaurus rex from Montana (MOR 1125, also known as B-rex). Medullary tissue is found only in female birds during ovulation, indicating that B-rex was of reproductive age. Further study indicates an age of 18 for this specimen. In 2016, it was finally confirmed by Mary Higby Schweitzer and Lindsay Zanno et al that the soft tissue within the Femur of MOR 1125 was medullary tissue. This also confirmed the identity of the specimen as a female. The discovery of medullary bone tissue within Tyrannosaurus may prove valuable in determining the sex of other dinosaur species in future examinations, as the chemical makeup of medullary tissue is unmistakable. Other tyrannosaurids exhibit extremely similar growth curves, although with lower growth rates corresponding to their lower adult sizes.
Over half of the known Tyrannosaurus rex specimens appear to have died within six years of reaching sexual maturity, a pattern which is also seen in other tyrannosaurs and in some large, long-lived birds and mammals today. These species are characterized by high infant mortality rates, followed by relatively low mortality among juveniles. Mortality increases again following sexual maturity, partly due to the stresses of reproduction. One study suggests that the rarity of juvenile Tyrannosaurus rex fossils is due in part to low juvenile mortality rates; the animals were not dying in large numbers at these ages, and so were not often fossilized. However, this rarity may also be due to the incompleteness of the fossil record or to the bias of fossil collectors towards larger, more spectacular specimens. In a 2013 lecture, Thomas Holtz Jr. suggested that dinosaurs "lived fast and died young" because they reproduced quickly whereas mammals have long life spans because they take longer to reproduce. Gregory S. Paul also writes that Tyrannosaurus reproduced quickly and died young, but attributes their short life spans to the dangerous lives they lived.
Sexual dimorphism
As the number of known specimens increased, scientists began to analyze the variation between individuals and discovered what appeared to be two distinct body types, or morphs, similar to some other theropod species. As one of these morphs was more solidly built, it was termed the 'robust' morph while the other was termed 'gracile'. Several morphological differences associated with the two morphs were used to analyze sexual dimorphism in Tyrannosaurus rex, with the 'robust' morph usually suggested to be female. For example, the pelvis of several 'robust' specimens seemed to be wider, perhaps to allow the passage of eggs. It was also thought that the 'robust' morphology correlated with a reduced chevron on the first tail vertebra, also ostensibly to allow eggs to pass out of the reproductive tract, as had been erroneously reported for crocodiles.
In recent years, evidence for sexual dimorphism has been weakened. A 2005 study reported that previous claims of sexual dimorphism in crocodile chevron anatomy were in error, casting doubt on the existence of similar dimorphism between Tyrannosaurus rex sexes. A full-sized chevron was discovered on the first tail vertebra of Sue, an extremely robust individual, indicating that this feature could not be used to differentiate the two morphs anyway. As Tyrannosaurus rex specimens have been found from Saskatchewan to New Mexico, differences between individuals may be indicative of geographic variation rather than sexual dimorphism. The differences could also be age-related, with 'robust' individuals being older animals.
Only a single Tyrannosaurus rex specimen has been conclusively shown to belong to a specific sex. Examination of B-rex demonstrated the preservation of soft tissue within several bones. Some of this tissue has been identified as a medullary tissue, a specialized tissue grown only in modern birds as a source of calcium for the production of eggshell during ovulation. As only female birds lay eggs, medullary tissue is only found naturally in females, although males are capable of producing it when injected with female reproductive hormones like estrogen. This strongly suggests that B-rex was female, and that she died during ovulation. Recent research has shown that medullary tissue is never found in crocodiles, which are thought to be the closest living relatives of dinosaurs, aside from birds. The shared presence of medullary tissue in birds and theropod dinosaurs is further evidence of the close evolutionary relationship between the two.
Posture
Modern representations in museums, art, and film show Tyrannosaurus rex with its body approximately parallel to the ground and tail extended behind the body to balance the head.
Like many bipedal dinosaurs, Tyrannosaurus rex was historically depicted as a 'living tripod', with the body at 45 degrees or less from the vertical and the tail dragging along the ground, similar to a kangaroo. This concept dates from Joseph Leidy's 1865 reconstruction of Hadrosaurus, the first to depict a dinosaur in a bipedal posture. In 1915, convinced that the creature stood upright, Henry Fairfield Osborn, former president of the American Museum of Natural History, further reinforced the notion in unveiling the first complete Tyrannosaurus rex skeleton arranged this way. It stood in an upright pose for 77 years, until it was dismantled in 1992.
By 1970, scientists realized this pose was incorrect and could not have been maintained by a living animal, as it would have resulted in the dislocation or weakening of several joints, including the hips and the articulation between the head and the spinal column. The inaccurate AMNH mount inspired similar depictions in many films and paintings (such as Rudolph Zallinger's famous mural The Age of Reptiles in Yale University's Peabody Museum of Natural History) until the 1990s, when films such as Jurassic Park introduced a more accurate posture to the general public.
Arms
When Tyrannosaurus rex was first discovered, the humerus was the only element of the forelimb known. For the initial mounted skeleton as seen by the public in 1915, Osborn substituted longer, three-fingered forelimbs like those of Allosaurus. However, a year earlier, Lawrence Lambe described the short, two-fingered forelimbs of the closely related Gorgosaurus. This strongly suggested that Tyrannosaurus rex had similar forelimbs, but this hypothesis was not confirmed until the first complete Tyrannosaurus rex forelimbs were identified in 1989, belonging to MOR 555 (the "Wankel rex"). The remains of Sue also include complete forelimbs. Tyrannosaurus rex arms are very small relative to overall body size, measuring only 1 meter (3.3 ft) long, and some scholars have labelled them as vestigial. However, the bones show large areas for muscle attachment, indicating considerable strength. This was recognized as early as 1906 by Osborn, who speculated that the forelimbs may have been used to grasp a mate during copulation. It has also been suggested that the forelimbs were used to assist the animal in rising from a prone position.
Another possibility is that the forelimbs held struggling prey while it was killed by the tyrannosaur's enormous jaws. This hypothesis may be supported by biomechanical analysis. Tyrannosaurus rex forelimb bones exhibit extremely thick cortical bone, which have been interpreted as evidence that they were developed to withstand heavy loads. The biceps brachii muscle of a full-grown Tyrannosaurus rex was capable of lifting 199 kilograms (439 lb) by itself; other muscles such as the brachialis would work along with the biceps to make elbow flexion even more powerful. The M. biceps muscle of T. rex was 3.5 times as powerful as the human equivalent. A Tyrannosaurus rex forearm had a limited range of motion, with the shoulder and elbow joints allowing only 40 and 45 degrees of motion, respectively. In contrast, the same two joints in Deinonychus allow up to 88 and 130 degrees of motion, respectively, while a human arm can rotate 360 degrees at the shoulder and move through 165 degrees at the elbow. The heavy build of the arm bones, strength of the muscles, and limited range of motion may indicate a system evolved to hold fast despite the stresses of a struggling prey animal. In the first detailed scientific description of Tyrannosaurus forelimbs, paleontologists Kenneth Carpenter and Matt Smith dismissed notions that the forelimbs were useless or that Tyrannosaurus rex was an obligate scavenger.
Soft tissue
In the March 2005 issue of Science, Mary Higby Schweitzer of North Carolina State University and colleagues announced the recovery of soft tissue from the marrow cavity of a fossilized leg bone from a Tyrannosaurus rex. The bone had been intentionally, though reluctantly, broken for shipping and then not preserved in the normal manner, specifically because Schweitzer was hoping to test it for soft tissue. Designated as the Museum of the Rockies specimen 1125, or MOR 1125, the dinosaur was previously excavated from the Hell Creek Formation. Flexible, bifurcating blood vessels and fibrous but elastic bone matrix tissue were recognized. In addition, microstructures resembling blood cells were found inside the matrix and vessels. The structures bear resemblance to ostrich blood cells and vessels. Whether an unknown process, distinct from normal fossilization, preserved the material, or the material is original, the researchers do not know, and they are careful not to make any claims about preservation. If it is found to be original material, any surviving proteins may be used as a means of indirectly guessing some of the DNA content of the dinosaurs involved, because each protein is typically created by a specific gene. The absence of previous finds may be the result of people assuming preserved tissue was impossible, therefore not looking. Since the first, two more tyrannosaurs and a hadrosaur have also been found to have such tissue-like structures. Research on some of the tissues involved has suggested that birds are closer relatives to tyrannosaurs than other modern animals.
In studies reported in Science in April 2007, Asara and colleagues concluded that seven traces of collagen proteins detected in purified Tyrannosaurus rex bone most closely match those reported in chickens, followed by frogs and newts. The discovery of proteins from a creature tens of millions of years old, along with similar traces the team found in a mastodon bone at least 160,000 years old, upends the conventional view of fossils and may shift paleontologists' focus from bone hunting to biochemistry. Until these finds, most scientists presumed that fossilization replaced all living tissue with inert minerals. Paleontologist Hans Larsson of McGill University in Montreal, who was not part of the studies, called the finds "a milestone", and suggested that dinosaurs could "enter the field of molecular biology and really slingshot paleontology into the modern world".
Subsequent studies in April 2008 confirmed the close connection of Tyrannosaurus rex to modern birds. Postdoctoral biology researcher Chris Organ at Harvard University announced, "With more data, they would probably be able to place T. rex on the evolutionary tree between alligators and chickens and ostriches." Co-author John M. Asara added, "We also show that it groups better with birds than modern reptiles, such as alligators and green anole lizards."
The presumed soft tissue was called into question by Thomas Kaye of the University of Washington and his co-authors in 2008. They contend that what was really inside the tyrannosaur bone was slimy biofilm created by bacteria that coated the voids once occupied by blood vessels and cells. The researchers found that what previously had been identified as remnants of blood cells, because of the presence of iron, were actually framboids, microscopic mineral spheres bearing iron. They found similar spheres in a variety of other fossils from various periods, including an ammonite. In the ammonite they found the spheres in a place where the iron they contain could not have had any relationship to the presence of blood. However, Schweitzer has strongly criticized Kaye's claims and argues that there's no reported evidence that biofilms can produce branching, hollow tubes like those noted in her study. San Antonio, Schweitzer and colleagues published an analysis in 2011 of what parts of the collagen had been recovered, finding that it was the inner parts of the collagen coil that had been preserved, as would have been expected from a long period of protein degradation. Other research challenges the identification of soft tissue as biofilm and confirms finding "branching, vessel-like structures" from within fossilized bone.
Thermoregulation
As of 2014, it is not clear if Tyrannosaurus was endothermic (warm-blooded). Tyrannosaurus, like most dinosaurs, was long thought to have an ectothermic ("cold-blooded") reptilian metabolism. The idea of dinosaur ectothermy was challenged by scientists like Robert T. Bakker and John Ostrom in the early years of the "Dinosaur Renaissance", beginning in the late 1960s. Tyrannosaurus rex itself was claimed to have been endothermic ("warm-blooded"), implying a very active lifestyle. Since then, several paleontologists have sought to determine the ability of Tyrannosaurus to regulate its body temperature. Histological evidence of high growth rates in young Tyrannosaurus rex, comparable to those of mammals and birds, may support the hypothesis of a high metabolism. Growth curves indicate that, as in mammals and birds, Tyrannosaurus rex growth was limited mostly to immature animals, rather than the indeterminate growth seen in most other vertebrates.
Oxygen isotope ratios in fossilized bone are sometimes used to determine the temperature at which the bone was deposited, as the ratio between certain isotopes correlates with temperature. In one specimen, the isotope ratios in bones from different parts of the body indicated a temperature difference of no more than 4 to 5 °C (7 to 9 °F) between the vertebrae of the torso and the tibia of the lower leg. This small temperature range between the body core and the extremities was claimed by paleontologist Reese Barrick and geochemist William Showers to indicate that Tyrannosaurus rex maintained a constant internal body temperature (homeothermy) and that it enjoyed a metabolism somewhere between ectothermic reptiles and endothermic mammals. Other scientists have pointed out that the ratio of oxygen isotopes in the fossils today does not necessarily represent the same ratio in the distant past, and may have been altered during or after fossilization (diagenesis). Barrick and Showers have defended their conclusions in subsequent papers, finding similar results in another theropod dinosaur from a different continent and tens of millions of years earlier in time (Giganotosaurus). Ornithischian dinosaurs also showed evidence of homeothermy, while varanid lizards from the same formation did not. Even if Tyrannosaurus rex does exhibit evidence of homeothermy, it does not necessarily mean that it was endothermic. Such thermoregulation may also be explained by gigantothermy, as in some living sea turtles.
Footprints
Two isolated fossilized footprints have been tentatively assigned to Tyrannosaurus rex. The first was discovered at Philmont Scout Ranch, New Mexico, in 1983 by American geologist Charles Pillmore. Originally thought to belong to a hadrosaurid, examination of the footprint revealed a large 'heel' unknown in ornithopod dinosaur tracks, and traces of what may have been a hallux, the Dewclaw-like fourth digit of the tyrannosaur foot. The footprint was published as the ichnogenus Tyrannosauripus pillmorei in 1994, by Martin Lockley and Adrian Hunt. Lockley and Hunt suggested that it was very likely the track was made by a Tyrannosaurus rex, which would make it the first known footprint from this species. The track was made in what was once a vegetated wetland mud flat. It measures 83 centimeters (33 in) long by 71 centimeters (28 in) wide.
A second footprint that may have been made by a Tyrannosaurus was first reported in 2007 by British paleontologist Phil Manning, from the Hell Creek Formation of Montana. This second track measures 72 centimeters (28 in) long, shorter than the track described by Lockley and Hunt. Whether or not the track was made by Tyrannosaurus is unclear, though Tyrannosaurus and Nanotyrannus are the only large theropods known to have existed in the Hell Creek Formation.
A set of footprints in Glenrock, Wyoming dating to the Maastrichtian stage of the late Cretaceous and hailing from the Lance Formation were recently described by Scott Persons, Phil Currie et al. in January 2016, and are believed to belong to either a juvenile Tyrannosaurus rex or the dubious tyrannosaurid genus Nanotyrannus lancensis. From measurements and based on the positions of the footprints, the animal was believed to be traveling at a walking speed of around 2.8 to 5 miles per hour and was estimated to have a hip height of 1.56 m (5.1 ft) to 2.06 m (6.8 ft).
Locomotion
There are two main issues concerning the locomotory abilities of Tyrannosaurus: how well it could turn; and what its maximum straight-line speed was likely to have been. Both are relevant to the debate about whether it was a hunter or a scavenger.
Tyrannosaurus may have been slow to turn, possibly taking one to two seconds to turn only 45° — an amount that humans, being vertically oriented and tailless, can spin in a fraction of a second. The cause of the difficulty is rotational inertia, since much of Tyrannosaurus' mass was some distance from its center of gravity, like a human carrying a heavy timber horizontally — although it might have reduced the average distance by arching its back and tail and pulling its head and forelimbs close to its body, rather like the way ice skaters pull their arms closer in order to spin faster.
Scientists have produced a wide range of maximum speed estimates, mostly around 11 meters per second (40 km/h; 25 mph), but a few as low as 5–11 meters per second (18–40 km/h; 11–25 mph), and a few as high as 20 meters per second (72 km/h; 45 mph). Researchers have to rely on various estimating techniques because, while there are many tracks of very large theropods walking, so far none have been found of very large theropods running—and this absence may indicate that they did not run. Scientists who think that Tyrannosaurus was able to run point out that hollow bones and other features that would have lightened its body may have kept adult weight to a mere 4.5 metric tons (5.0 short tons) or so, or that other animals like ostriches and horses with long, flexible legs are able to achieve high speeds through slower but longer strides. Additionally, some have argued that Tyrannosaurus had relatively larger leg muscles than any animal alive today, which could have enabled fast running at 40–70 kilometers per hour (25–43 mph).
Jack Horner and Don Lessem argued in 1993 that Tyrannosaurus was slow and probably could not run (no airborne phase in mid-stride), because its ratio of femur (thigh bone) to Tibia (shin bone) length was greater than 1, as in most large theropods and like a modern elephant. However, Holtz (1998) noted that tyrannosaurids and some closely related groups had significantly longer distal hindlimb components (shin plus foot plus toes) relative to the femur length than most other theropods, and that tyrannosaurids and their close relatives had a tightly interlocked metatarsus that more effectively transmitted locomotory forces from the foot to the lower leg than in earlier theropods ("metatarsus" means the foot bones, which function as part of the leg in digitigrade animals). He therefore concluded that tyrannosaurids and their close relatives were the fastest large theropods. Thomas Holtz Jr. echoed these sentiments in his 2013 lecture, stating that the giant allosaurs had shorter feet for the same body size than Tyrannosaurus, whereas Tyrannosaurus had longer, skinnier and more interlocked feet for the same body size; attributes of faster moving animals.
A study by Eric Snively and Anthony P. Russel published in 2003 also found that the tyrannosaurid arctometatarsals and elastic ligaments worked together in what he called a 'tensile keystone model' to strengthen the feet of Tyrannosaurus, increase the animal's stability and add greater resistance to dissociation over that of other theropod families; while still allowing resiliency that is otherwise reduced in ratites, horses, giraffids and other animals with metapodia to a single element. The study also pointed out that elastic ligaments in larger vertebrates could store and return relatively more elastic strain energy, which could have improved locomotor efficiency and decrease the strain energy transferred to the bones. The study suggested that this mechanism could have worked efficiently in tyrannosaurids as well. Hence, the study involved identifying the type of ligaments attached to the metatarsals, then how they functioned together and comparing it to those of other theropods and modern day analogs. The scientists found that arctometatarsals may have enabled tyrannosaurid feet to absorb forces such as linear deceleration, lateral acceleration and torsion more effectively than those of other theropods. It is also stated in their study that this may imply, though not demonstrate, that tyrannosaurids such as Tyrannosaurus had greater agility than other large theropods without an arctometatarsus.
Christiansen (1998) estimated that the leg bones of Tyrannosaurus were not significantly stronger than those of elephants, which are relatively limited in their top speed and never actually run (there is no airborne phase), and hence proposed that the dinosaur's maximum speed would have been about 11 meters per second (40 km/h; 25 mph), which is about the speed of a human sprinter. But he also noted that such estimates depend on many dubious assumptions.
Farlow and colleagues (1995) have argued that a Tyrannosaurus weighing 5.4 metric tons (6.0 short tons) to 7.3 metric tons (8.0 short tons) would have been critically or even fatally injured if it had fallen while moving quickly, since its torso would have slammed into the ground at a deceleration of 6 g (six times the acceleration due to gravity, or about 60 meters/s²) and its tiny arms could not have reduced the impact. However, giraffes have been known to gallop at 50 kilometers per hour (31 mph), despite the risk that they might break a leg or worse, which can be fatal even in a "safe" environment such as a zoo. Thus it is possible that Tyrannosaurus also moved fast when necessary and had to accept such risks.
In another study, Gregory S. Paul pointed out that the flexed kneed and digitigrade adult Tyrannosaurus were much better adapted for running than elephants or humans, pointing out that Tyrannosaurus had a large ilium bone and cnemial crest that would have supported large muscles needed for running. He also mentioned that Alexander's (1989) formula to calculate speed by bone strength was only partly reliable. He suggests that the formula is overly sensitive to bone length; making long bones artificially weak. He also pointed out that the lowered risk of being wounded in combat may have been worth the risk of Tyrannosaurus falling while running.
Most recent research on Tyrannosaurus locomotion does not support speeds faster than 40 kilometers per hour (25 mph), i.e. moderate-speed running. For example, a 2002 paper in Nature used a mathematical model (validated by applying it to three living animals, alligators, chickens, and humans; later eight more species including emus and ostriches) to gauge the leg muscle mass needed for fast running (over 40 km/h or 25 mph). They found that proposed top speeds in excess of 40 kilometers per hour (25 mph) were infeasible, because they would require very large leg muscles (more than approximately 40–86% of total body mass). Even moderately fast speeds would have required large leg muscles. This discussion is difficult to resolve, as it is unknown how large the leg muscles actually were in Tyrannosaurus. If they were smaller, only 18 kilometers per hour (11 mph) walking or jogging might have been possible.
A study in 2007 used computer models to estimate running speeds, based on data taken directly from fossils, and claimed that Tyrannosaurus rex had a top running speed of 8 meters per second (29 km/h; 18 mph). An average professional football (soccer) player would be slightly slower, while a human sprinter can reach 12 meters per second (43 km/h; 27 mph). These computer models predict a top speed of 17.8 meters per second (64 km/h; 40 mph) for a 3-kilogram (6.6 lb) Compsognathus (probably a juvenile individual).
However, in 2010, Scott Persons, a graduate student from the University of Alberta proposed that Tyrannosaurus's speed may have been enhanced by strong tail muscles. He found that theropods such as T rex had certain muscle arrangements that are different from modern day birds and mammals but with some similarities to modern reptiles. He concluded that the caudofemoralis muscles which link the tail bones and the upper leg bones could have assisted Tyrannosaurus in leg retraction and enhanced its running ability, agility and balance. The caudofemoralis muscle would have been a key muscle in femoral retraction; pulling back the leg at the femur. The study also found that theropod skeletons such as those of Tyrannosaurus had adaptations (such as elevated transverse processes in the tail vertebrae) to enable the growth of larger tail muscles and that Tyrannosaurus's tail muscle mass may have been underestimated by over 25 percent and perhaps as much as 45 percent. The caudofemoralis muscle was found to comprise 58 percent of the muscle mass in the tail of Tyrannosaurus. Tyrannosaurus also had the largest absolute and relative caudofemoralis muscle mass out of the three extinct organisms in the study. This is because Tyrannosaurus also had additional adaptations to enable large tail muscles; the elongation of its tail's hemal arches. According to Persons, the increase in tail muscle mass would have moved the center of mass closer to the hindquarters and hips which would have lessened the strain on the leg muscles to support its weight; improving its overall balance and agility. This would also have made the animal less front-heavy, thus reducing rotational inertia. Persons also notes that the tail is also rich in tendons and septa which could have been stores of elastic energy, and thereby improved locomotive efficiency. Persons adds that this means non-avian theropods actually had broader tails than previously depicted, as broad or broader laterally than dorsoventrally near the base.
Heinrich Mallison from Berlin's Museum of Natural History also presented a theory in 2011, suggesting that Tyrannosaurus and many other dinosaurs may have achieved relatively high speeds through short rapid strides instead of the long strides employed by modern birds and mammals when running, likening their movement to power-walking. This, according to Mallison, would have been achievable irrespective of joint strength and lessened the need for additional muscle mass in the legs, particularly at the ankles. To support his theory, Mallison assessed the limbs of various dinosaurs and found that they were different from those of modern mammals and birds; having their stride length greatly limited by their skeletons, but also having relatively large muscles at the hindquarters. However, he found a few similarities between the muscles in dinosaurs and race-walkers; having less muscle mass in the ankles but more at the hindquarters. Mallison suggests that the differences between dinosaurs, extant mammals, and birds would also have made equations to calculate speed from stride length inapplicable to dinosaurs. John Hutchinson however advised caution regarding this theory, suggesting that they must first look into dinosaur muscles to see how frequently they could have contracted.
Those who argue that Tyrannosaurus was incapable of running estimate the top speed of Tyrannosaurus at about 17 kilometers per hour (11 mph). This is still faster than its most likely prey species, hadrosaurids and ceratopsians. In addition, some advocates of the idea that Tyrannosaurus was a predator claim that tyrannosaur running speed is not important, since it may have been slow but still faster than its probable prey. Thomas Holtz also noted that Tyrannosaurus had proportionately longer feet than the animals it hunted: duck-billed dinosaurs and horned dinosaurs. However, Paul and Christiansen (2000) argued that at least the later ceratopsians had upright forelimbs and the larger species may have been as fast as rhinos. Healed Tyrannosaurus bite wounds on ceratopsian fossils are interpreted as evidence of attacks on living ceratopsians (see below). If the ceratopsians that lived alongside Tyrannosaurus were fast, that casts doubt on the argument that Tyrannosaurus did not have to be fast to catch its prey.
Brain and senses
A study conducted by Lawrence Witmer and Ryan Ridgely of Ohio University found that Tyrannosaurus shared the heightened sensory abilities of other coelurosaurs, highlighting relatively rapid and coordinated eye and head movements, as well as an enhanced ability to sense low frequency sounds that would allow tyrannosaurs to track prey movements from long distances and an enhanced sense of smell. A study published by Kent Stevens of the University of Oregon concluded that Tyrannosaurus had keen vision. By applying modified perimetry to facial reconstructions of several dinosaurs including Tyrannosaurus, the study found that Tyrannosaurus had a binocular range of 55 degrees, surpassing that of modern hawks, and had 13 times the visual acuity of a human, thereby surpassing the visual acuity of an eagle which is only 3.6 times that of a person. This would have allowed Tyrannosaurus to discern objects as far as 6 km (3.7 mi) away, which is greater than the 1.6 km (1 mi) that a human can see.
Thomas Holtz Jr. would note that high depth perception of Tyrannosaurus may have been due to the prey it had to hunt; noting that it had to hunt horned dinosaurs such as Triceratops, armored dinosaurs such as Ankylosaurus and the duck-billed dinosaurs may have had complex social behaviors. He would suggest that this made precision more crucial for Tyrannosaurus enabling it to, "get in, get that blow in and take it down." In contrast, Acrocanthosaurus had limited depth perception because they hunted large sauropods, which were relatively rare during the time of Tyrannosaurus.
Tyrannosaurus had very large olfactory bulbs and olfactory nerves relative to their brain size, the organs responsible for a heightened sense of smell. This suggests that the sense of smell was highly developed, and implies that tyrannosaurs could detect carcasses by scent alone across great distances. The sense of smell in tyrannosaurs may have been comparable to modern vultures, which use scent to track carcasses for scavenging. Research on the olfactory bulbs has shown that Tyrannosaurus rex had the most highly developed sense of smell of 21 sampled non-avian dinosaur species.
Somewhat unusually among theropods, T. rex had a very long cochlea. The length of the cochlea is often related to hearing acuity, or at least the importance of hearing in behavior, implying that hearing was a particularly important sense to tyrannosaurs. Specifically, data suggests that Tyrannosaurus rex heard best in the low-frequency range, and that low-frequency sounds were an important part of tyrannosaur behavior.
A study by Grant R. Hurlburt, Ryan C. Ridgely and Lawrence Witmer would obtain estimates for Encephalization Quotients (EQs), based on reptiles and birds, as well as estimates for the ratio of cerebrum to brain mass. The study would conclude that Tyrannosaurus had the relatively largest brain of all adult non-avian dinosaurs with the exception of certain small maniraptoriforms (Bambiraptor, Troodon and Ornithomimus). The study would however find that Tyrannosaurus's relative brain size was still within the range of modern reptiles, being at most 2 standard deviations above the mean of non-avian reptile EQs. The estimates for the ratio of cerebrum mass to brain mass would range from 47.5 to 49.53 percent. According to the study, this is more than the lowest estimates for extant birds (44.6 percent), but still close to the typical ratios of the smallest sexually mature alligators which range from 45.9–47.9 percent.
Feeding strategies
A 2012 study by scientists Karl Bates and Peter Falkingham suggested that the bite force of Tyrannosaurus could have been the strongest of any terrestrial animal that has ever lived. The calculations suggested that adult T. rex could have generated from 35,000 to 57,000 Newtons of force in the back teeth. However, even higher estimates were made by professor Mason B. Meers of the University of Tampa in 2003. In his study, Meers estimated a possible bite force of around 183,000 to 235,000 Newtons or 18.3 to 23.5 metric tons (20.2 to 25.9 short tons). Research done by Stephan Lautenschlager et al. of the University of Bristol, also reveals Tyrannosaurus was also capable of a maximum jaw gape of around 63.5 degrees, a necessary adaptation for a wide range of jaw angles in order to power the creature's strong bite.
The debate about whether Tyrannosaurus was a predator or a pure scavenger is as old as the debate about its locomotion. Lambe (1917) described a good skeleton of Tyrannosaurus close relative Gorgosaurus and concluded that it and therefore also Tyrannosaurus was a pure scavenger, because the Gorgosaurus teeth showed hardly any wear. This argument is no longer taken seriously, because theropods replaced their teeth quite rapidly. Ever since the first discovery of Tyrannosaurus most scientists have speculated that it was a predator; like modern large predators it would readily scavenge or steal another predator's kill if it had the opportunity.
Paleontologist Jack Horner has been a major advocate of the idea that Tyrannosaurus was exclusively a scavenger and did not engage in active hunting at all, though Horner himself has claimed that he never published this idea in the peer-reviewed scientific literature and used it mainly as a tool to teach a popular audience, particularly children, the dangers of making assumptions in science (such as assuming T. rex was a hunter) without using evidence. Nevertheless, Horner presented several arguments in the popular literature to support the pure scavenger hypothesis:
Other evidence suggests hunting behavior in Tyrannosaurus. The eye sockets of tyrannosaurs are positioned so that the eyes would point forward, giving them binocular vision slightly better than that of modern hawks. Horner also pointed out that the tyrannosaur lineage had a history of steadily improving binocular vision. It is not obvious why natural selection would have favored this long-term trend if tyrannosaurs had been pure scavengers, which would not have needed the advanced depth perception that stereoscopic vision provides. In modern animals, binocular vision is found mainly in predators.
A skeleton of the hadrosaurid Edmontosaurus annectens has been described from Montana with healed tyrannosaur-inflicted damage on its tail vertebrae. The fact that the damage seems to have healed suggests that the Edmontosaurus survived a tyrannosaur's attack on a living target, i.e. the tyrannosaur had attempted active predation. There is also evidence for an aggressive interaction between a Triceratops and a Tyrannosaurus in the form of partially healed tyrannosaur tooth marks on a Triceratops brow horn and squamosal (a bone of the neck frill); the bitten horn is also broken, with new bone growth after the break. It is not known what the exact nature of the interaction was, though: either animal could have been the aggressor. Since the Triceratops wounds healed, it is most likely that the Triceratops survived the encounter and managed to overcome the Tyrannosaurus. Paleontologist Peter Dodson estimates that in a battle against a bull Triceratops, the Triceratops had the upper hand and would successfully defend itself by inflicting fatal wounds to the Tyrannosaurus using its sharp horns.
When examining Sue, paleontologist Pete Larson found a broken and healed fibula and tail vertebrae, scarred facial bones and a tooth from another Tyrannosaurus embedded in a neck vertebra. If correct, these might be strong evidence for aggressive behavior between tyrannosaurs but whether it would have been competition for food and mates or active cannibalism is unclear. However, further recent investigation of these purported wounds has shown that most are infections rather than injuries (or simply damage to the fossil after death) and the few injuries are too general to be indicative of intraspecific conflict. Some researchers argue that if Tyrannosaurus were a scavenger, another dinosaur had to be the top predator in the Amerasian Upper Cretaceous. Top prey were the larger marginocephalians and ornithopods. The other tyrannosaurids share so many characteristics that only small dromaeosaurs and troodontids remain as feasible top predators. In this light, scavenger hypothesis adherents have suggested that the size and power of tyrannosaurs allowed them to steal kills from smaller predators, although they may have had a hard time finding enough meat to scavenge, being outnumbered by smaller theropods. Most paleontologists accept that Tyrannosaurus was both an active predator and a scavenger like most large carnivores.
Tyrannosaurus may have had infectious saliva used to kill its prey. This theory was first proposed by William Abler. Abler examined the teeth of tyrannosaurids between each tooth serration; the serrations may have held pieces of carcass with bacteria, giving Tyrannosaurus a deadly, infectious bite much like the Komodo dragon was thought to have. However, Jack Horner regards Tyrannosaurus tooth serrations as more like cubes in shape than the serrations on a Komodo monitor's teeth, which are rounded. All forms of saliva contain possibly hazardous bacteria, so the prospect of it being used as a method of predation is disputable.
Tyrannosaurus, and most other theropods, probably primarily processed carcasses with lateral shakes of the head, like crocodilians. The head was not as maneuverable as the skulls of allosauroids, due to flat joints of the neck vertebrae.
Cannibalism
A study from Currie, Horner, Erickson and Longrich in 2010 has been put forward as evidence of cannibalism in the genus Tyrannosaurus. They studied some Tyrannosaurus specimens with tooth marks in the bones, attributable to the same genus. The tooth marks were identified in the humerus, foot bones and metatarsals, and this was seen as evidence for opportunistic scavenging, rather than wounds caused by intraspecific combat. In a fight, they proposed it would be difficult to reach down to bite in the feet of a rival, making it more likely that the bite marks were made in a carcass. As the bite marks were made in body parts with relatively scanty amounts of flesh, it is suggested that the Tyrannosaurus was feeding on a carcass in which the more fleshy parts had already been consumed. They were also open to the possibility that other tyrannosaurids practiced cannibalism. Other evidence for cannibalism has been unearthed.
Pack behavior
Philip J. Currie of the University of Alberta has suggested that Tyrannosaurus may have been pack animals. Currie compared Tyrannosaurus rex favorably to related species Tarbosaurus bataar and Albertosaurus sarcophagus, fossil evidence from which Currie had previously used to suggest that they lived in packs. Currie pointed out that a find in South Dakota preserved three Tyrannosaurus rex skeletons in close proximity to each other. After using CT scanning, Currie stated that Tyrannosaurus would have been capable of such complex behavior, because its brain size is three times greater than what would be expected for an animal of its size. Currie elaborated that Tyrannosaurus had a larger brain-to-body-size proportion than crocodiles and three times more than plant eating dinosaurs such as Triceratops of the same size. Currie believed Tyrannosaurus to be six times smarter than most dinosaurs and other reptiles. Because the available prey, such as Triceratops and Ankylosaurus, were well-armored, and that others were fast-moving, it would have been necessary for Tyrannosaurus to hunt in groups. Currie speculated that juveniles and adults would have hunted together, with the faster juveniles chasing down the prey and the more powerful adults making the kill, by analogy to modern-day pack hunters where each member contributes a skill.
However, Currie's pack-hunting hypothesis has been harshly criticized by other scientists. Brian Switek, writing for The Guardian in 2011, noted that Currie's pack hypothesis has not been presented as research in a peer-reviewed scientific journal, but primarily in relation to a television special and tie-in book called Dino Gangs. Switek also noted that Currie's argument for pack hunting in Tyrannosaurus rex is primarily based on analogy to a different species, Tarbosaurus bataar, and that the supposed evidence for pack hunting in T. bataar itself has not yet been published and subjected to scientific scrutiny. According to Switek and other scientists who have participated in panel discussions about the Dino Gangs television program, the evidence for pack hunting in Tarbosaurus and Albertosaurus is weak, based primarily on the association of several skeletons, for which numerous alternative explanations have been proposed (e.g. drought or floods forcing numerous specimens together to die in one place). In fact, Switek notes that the Albertosaurus bonebed site, on which Currie has based most of the interpretations of supposed pack hunting in related species, preserves geological evidence of just such a flood. Switek said, "bones alone are not enough to reconstruct dinosaur behaviour. The geological context in which those bones are found – the intricate details of ancient environments and the pace of prehistoric time – are essential to investigating the lives and deaths of dinosaurs," and noted that Currie must first describe the geological evidence from other tyrannosaur bonebed sites before jumping to conclusions about social behavior. Switek described the sensational claims provided in press releases and news stories surrounding the Dino Gangs program as "nauseating hype" and noted that the production company responsible for the program, Atlantic Productions, has a poor record involving exaggerating claims about new fossil discoveries, most notably the controversial claim it published regarding the supposed early human ancestor Darwinius, which soon turned out to be a relative of lemurs instead.
Lawrence Witmer pointed out that social behavior can't be determined by brain endocasts and the brains of solitary leopards are identical to those of a cooperatively hunting lion; estimated brain sizes only show that an animal may have hunted in groups. In his opinion, the brains of tyrannosaurs were large enough for what he dubs "communal hunting", a semi-organized behavior that falls between solitary and cooperative hunting. Witmer claims that communal hunting is a step towards the evolution of cooperative hunting. He found it hard to believe that tyrannosaurs wouldn't have exploited the opportunity to join others in making a kill, and thus decrease risk and increase their chances of success.
On July 23, 2014, evidence, for the first time, in the form of fossilized trackways in Canada, showed that tyrannosaurs may have hunted in groups.
Pathology
In 2001, Bruce Rothschild and others published a study examining evidence for stress fractures and tendon avulsions in theropod dinosaurs and the implications for their behavior. Since stress fractures are caused by repeated trauma rather than singular events they are more likely to be caused by regular behavior than other types of injuries. Of the 81 Tyrannosaurus foot bones examined in the study one was found to have a stress fracture, while none of the 10 hand bones were found to have stress fractures. The researchers found tendon avulsions only among Tyrannosaurus and Allosaurus. An avulsion injury left a divot on the humerus of Sue the T. rex, apparently located at the origin of the deltoid or teres major muscles. The presence of avulsion injuries being limited to the forelimb and shoulder in both Tyrannosaurus and Allosaurus suggests that theropods may have had a musculature more complex than and functionally different from those of birds. The researchers concluded that Sue's tendon avulsion was probably obtained from struggling prey. The presence of stress fractures and tendon avulsions in general provides evidence for a "very active" predation-based diet rather than obligate scavenging.
A 2009 study showed that holes in the skulls of several specimens that were previously explained by intraspecific attacks might have been caused by Trichomonas-like parasites that commonly infect avians. Further evidence of intraspecific attack would however be found by Joseph Peterson and his colleagues in the juvenile Tyrannosaurus nicknamed Jane. Peterson and his team found that Jane's skull showed healed puncture wounds on the upper jaw and snout which they believe came from another juvenile Tyrannosaurus. Subsequent CT scans of Jane's skull would further confirm the team's hypothesis, showing that the puncture wounds came from a traumatic injury and that there was subsequent healing. The team would also state that Jane's injuries were structurally different from the parasite-induced lesions found in Sue and that Jane's injuries were on her face whereas the parasite that infected Sue caused lesions to the lower jaw.
Paleoecology
Tyrannosaurus lived during what is referred to as the Lancian faunal stage (Maastrichtian age) at the end of the Late Cretaceous. Tyrannosaurus ranged from Canada in the north to at least Texas and New Mexico in the south of Laramidia. During this time Triceratops was the major herbivore in the northern portion of its range, while the titanosaurian sauropod Alamosaurus "dominated" its southern range. Tyrannosaurus remains have been discovered in different ecosystems, including inland and coastal subtropical, and semi-arid plains.
Several notable Tyrannosaurus remains have been found in the Hell Creek Formation. During the Maastrichtian this area was subtropical, with a warm and humid climate. The flora consisted mostly of angiosperms, but also included trees like dawn redwood (Metasequoia) and Araucaria. Tyrannosaurus shared this ecosystem with Triceratops, related ceratopsians Nedoceratops, Tatankaceratops and Torosaurus, the hadrosaurid Edmontosaurus annectens and possibly a species of Parasaurolophus, the armored dinosaurs Denversaurus, Edmontonia and Ankylosaurus, the dome headed dinosaurs Pachycephalosaurus, Stygimoloch, Sphaerotholus, and Dracorex, the hypsilophodont Thescelosaurus, and the theropods Ornithomimus, Struthiomimus, Orcomimus, Acheroraptor, Dakotaraptor, Richardoestesia, Paronychodon, Pectinodon and Troodon.
Another formation with tyrannosaur remains is the Lance Formation of Wyoming. This has been interpreted as a bayou environment similar to today's Gulf Coast. The fauna was very similar to Hell Creek, but with Struthiomimus replacing its relative Ornithomimus. The small ceratopsian Leptoceratops also lived in the area.
In its southern range Tyrannosaurus lived alongside the titanosaur Alamosaurus, the ceratopsians Torosaurus, Bravoceratops and Ojoceratops, hadrosaurs which consisted of a species of Edmontosaurus, Kritosaurus and a possible species of Gryposaurus, the nodosaur Glyptodontopelta, the oviraptorid Ojoraptosaurus, possible species of the theropods Troodon and Richardoestesia, and the pterosaur Quetzalcoatlus. The region is thought to have been dominated by semi-arid inland plains, following the probable retreat of the Western Interior Seaway as global sea levels fell.
Tyrannosaurus may have also inhabited Mexico's Lomas Coloradas formation in Sonora. Though skeletal evidence is lacking, six shed and broken teeth from the fossil bed have been thoroughly compared with other theropod genera and appear to be identical to those of Tyrannosaurus. If true, the evidence indicates the range of Tyrannosaurus was possibly more extensive than previously believed. It is possible that tyrannosaurs were originally Asian species, migrating to North America before the end of the Cretaceous period.
History of research
Henry Fairfield Osborn, president of the American Museum of Natural History, named Tyrannosaurus rex in 1905. The generic name is derived from the Greek words τυράννος (tyrannos, meaning "tyrant") and σαύρος (sauros, meaning "lizard"). Osborn used the Latin word rex, meaning "king", for the specific name. The full binomial therefore translates to "tyrant lizard the king" or "King Tyrant Lizard", emphasizing the animal's size and perceived dominance over other species of the time.
Earliest finds
Teeth from what is now documented as a Tyrannosaurus rex were found in 1874 by Arthur Lakes near Golden, Colorado. In the early 1890s, John Bell Hatcher collected postcranial elements in eastern Wyoming. The fossils were believed to be from a large species of Ornithomimus (O. grandis) but are now considered Tyrannosaurus rex remains. Vertebral fragments found by Edward Drinker Cope in western South Dakota in 1892 and assigned to Manospondylus gigas have also been recognized as belonging to Tyrannosaurus rex.
Barnum Brown, assistant curator of the American Museum of Natural History, found the first partial skeleton of Tyrannosaurus rex in eastern Wyoming in 1900. H. F. Osborn originally named this skeleton Dynamosaurus imperiosus in a paper in 1905. Brown found another partial skeleton in the Hell Creek Formation in Montana in 1902. Osborn used this holotype to describe Tyrannosaurus rex in the same paper in which D. imperiosus was described. In 1906, Osborn recognized the two as Synonyms, and acted as first revisor by selecting Tyrannosaurus as the valid name. The original Dynamosaurus material resides in the collections of the Natural History Museum, London.
In total, Brown found five Tyrannosaurus partial skeletons. In 1941, Brown's 1902 find was sold to the Carnegie Museum of Natural History in Pittsburgh, Pennsylvania. Brown's fourth and largest find, also from Hell Creek, is on display in the American Museum of Natural History in New York.
Manospondylus
The first named fossil specimen which can be attributed to Tyrannosaurus rex consists of two partial vertebrae (one of which has been lost) found by Edward Drinker Cope in 1892. Cope believed that they belonged to an "agathaumid" (ceratopsid) dinosaur, and named them Manospondylus gigas, meaning "giant porous vertebra" in reference to the numerous openings for blood vessels he found in the bone. The M. gigas remains were later identified as those of a theropod rather than a ceratopsid, and H.F. Osborn recognized the similarity between M. gigas and Tyrannosaurus rex as early as 1917. However, owing to the fragmentary nature of the Manospondylus vertebrae, Osborn did not synonymize the two genera.
In June 2000, the Black Hills Institute located the type locality of M. gigas in South Dakota and unearthed more tyrannosaur bones there. These were judged to represent further remains of the same individual, and to be identical to those of Tyrannosaurus rex. According to the rules of the International Code of Zoological Nomenclature (ICZN), the system that governs the scientific naming of animals, Manospondylus gigas should therefore have priority over Tyrannosaurus rex, because it was named first. However, the Fourth Edition of the ICZN, which took effect on January 1, 2000, states that "the prevailing usage must be maintained" when "the senior synonym or homonym has not been used as a valid name after 1899" and "the junior synonym or homonym has been used for a particular taxon, as its presumed valid name, in at least 25 works, published by at least 10 authors in the immediately preceding 50 years ..." Tyrannosaurus rex may qualify as the valid name under these conditions and would most likely be considered a nomen protectum ("protected name") under the ICZN if it is ever formally published on, which it has not yet been. Manospondylus gigas could then be deemed a nomen oblitum ("forgotten name").
Notable specimens
Sue Hendrickson, amateur paleontologist, discovered the most complete (approximately 85%) and the largest Tyrannosaurus fossil skeleton known in the Hell Creek Formation near Faith, South Dakota, on August 12, 1990. This Tyrannosaurus, nicknamed Sue in her honor, was the object of a legal battle over its ownership. In 1997 this was settled in favor of Maurice Williams, the original land owner. The fossil collection was purchased by the Field Museum of Natural History at auction for $7.6 million, making it the most expensive dinosaur skeleton to date. From 1998 to 1999 Field Museum of Natural History preparators spent over 25,000 man-hours taking the rock off each of the bones. The bones were then shipped off to New Jersey where the mount was made. The finished mount was then taken apart, and along with the bones, shipped back to Chicago for the final assembly. The mounted skeleton opened to the public on May 17, 2000 in the great hall (Stanley Field Hall) at the Field Museum of Natural History. A study of this specimen's fossilized bones showed that Sue reached full size at age 19 and died at age 28, the longest any tyrannosaur is known to have lived. Early speculation that Sue may have died from a bite to the back of the head was not confirmed. Though subsequent study showed many pathologies in the skeleton, no bite marks were found. Damage to the back of the skull may have been caused by post-mortem trampling. Recent speculation indicates that Sue may have died of starvation after contracting a parasitic infection from eating diseased meat; the resulting infection would have caused inflammation in the throat, ultimately leading Sue to starve because she could no longer swallow food. This hypothesis is substantiated by smooth-edged holes in her skull which are similar to those caused in modern-day birds that contract the same parasite.
Another Tyrannosaurus, nicknamed Stan, in honor of amateur paleontologist Stan Sacrison, was found in the Hell Creek Formation near Buffalo, South Dakota, in the spring of 1987. It was not collected until 1992, as it was mistakenly thought to be a Triceratops skeleton. Stan is 63% complete and is on display in the Black Hills Institute of Geological Research in Hill City, South Dakota, after an extensive world tour during 1995 and 1996. This tyrannosaur, too, was found to have many bone pathologies, including broken and healed ribs, a broken (and healed) neck and a spectacular hole in the back of its head, about the size of a Tyrannosaurus tooth.
In the summer of 2000, Jack Horner discovered five Tyrannosaurus skeletons near the Fort Peck Reservoir in Montana. One of the specimens was reported to be perhaps the largest Tyrannosaurus ever found.
In 2001, a 50% complete skeleton of a juvenile Tyrannosaurus was discovered in the Hell Creek Formation in Montana, by a crew from the Burpee Museum of Natural History of Rockford, Illinois. Dubbed Jane, the find was initially considered the first known skeleton of the pygmy tyrannosaurid Nanotyrannus but subsequent research has revealed that it is more likely a juvenile Tyrannosaurus. It is the most complete and best preserved juvenile example known to date. Jane has been examined by Jack Horner, Pete Larson, Robert Bakker, Greg Erickson, and several other renowned paleontologists, because of the uniqueness of her age. Jane is currently on exhibit at the Burpee Museum of Natural History in Rockford, Illinois.
In a press release on April 7, 2006, Bozeman Campus, Montana State University, USA revealed that it possessed the largest Tyrannosaurus skull yet discovered. Discovered in the 1960s and only recently reconstructed, the skull measured 59 inches (150 cm) long compared to the 55.4 inches (141 cm) of Sue's skull, a difference of 6.5%. However, it had been incorrectly reconstructed, and has since been corrected to 134 cm (53 in).
In popular culture
Since it was first described in 1905, Tyrannosaurus rex has become the most widely recognized dinosaur species in popular culture. It is the only dinosaur that is commonly known to the general public by its full scientific name (binomial name) (Tyrannosaurus rex), and the scientific abbreviation T. rex has also come into wide usage. Robert T. Bakker notes this in The Dinosaur Heresies and explains that a name like "Tyrannosaurus rex is just irresistible to the tongue."