
Latin Astrocytus Dorlands/Elsevier 12165688 | NeuroLex ID Astrocyte | |
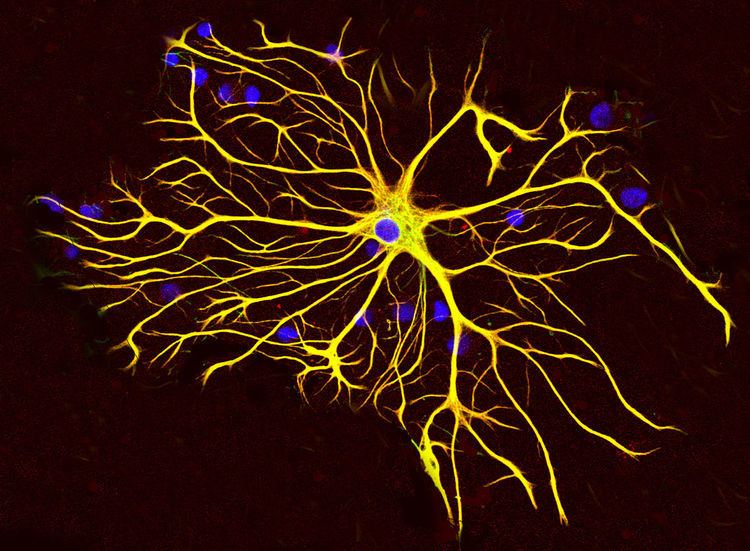 | ||
Astrocytes (Astro from Greek astron = star and cyte from Greek "kyttaron" = cell), also known collectively as astroglia, are characteristic star-shaped glial cells in the brain and spinal cord. The proportion of astrocytes in the brain is not well defined. Depending on the counting technique used, studies have found that the astrocyte proportion varies by region and ranges from 20% to 40% of all glia. They perform many functions, including biochemical support of endothelial cells that form the blood–brain barrier, provision of nutrients to the nervous tissue, maintenance of extracellular ion balance, and a role in the repair and scarring process of the brain and spinal cord following traumatic injuries.
Contents
- Structure
- Development
- Function
- Bergmann glia
- Tripartite synapse
- Astrocytomas
- Neurodevelopment disorders
- Chronic pain
- Two states of persistent pain
- Other pathologies
- Recent studies
- Classification
- Lineage and antigenic phenotype
- Anatomical classification
- Transporterreceptor classification
- References
Research since the mid-1990s has shown that astrocytes propagate intercellular Ca2+ waves over long distances in response to stimulation, and, similar to neurons, release transmitters (called gliotransmitters) in a Ca2+-dependent manner. Data suggest that astrocytes also signal to neurons through Ca2+-dependent release of glutamate. Such discoveries have made astrocytes an important area of research within the field of neuroscience.
Structure
Astrocytes are a sub-type of glial cells in the central nervous system. They are also known as astrocytic glial cells. Star-shaped, their many processes envelop synapses made by neurons. Astrocytes are classically identified using histological analysis; many of these cells express the intermediate filament glial fibrillary acidic protein (GFAP). Several forms of astrocytes exist in the central nervous system including fibrous (in white matter), protoplasmic (in grey matter), and radial. The fibrous glia are usually located within white matter, have relatively few organelles, and exhibit long unbranched cellular processes. This type often has "vascular feet" that physically connect the cells to the outside of capillary walls when they are in proximity to them. The protoplasmic glia are the most prevalent and are found in grey matter tissue, possess a larger quantity of organelles, and exhibit short and highly branched tertiary processes. The radial glia are disposed in planes perpendicular to the axes of ventricles. One of their processes abuts the pia mater, while the other is deeply buried in gray matter. Radial glia are mostly present during development, playing a role in neuron migration. Müller cells of the retina and Bergmann glia cells of the cerebellar cortex represent an exception, being present still during adulthood. When in proximity to the pia mater, all three forms of astrocytes send out processes to form the pia-glial membrane.
Development
Astrocytes are macroglial cells in the central nervous system. Astrocytes are derived from heterogeneous populations of progenitor cells in the neuroepithelium of the developing central nervous system. Recent works, summarized in a review by Rowitch and Kriegstein, indicate that there is remarkable similarity between the well known genetic mechanisms that specify the lineage of diverse neuron subtypes and that of macroglial cells. Just as with neuronal cell specification, canonical signaling factors like Sonic hedgehog (SHH), Fibroblast growth factor (FGFs), WNTs and bone morphogenetic proteins (BMPs), provide positional information to developing macroglial cells through morphogen gradients along the dorsal–ventral, anterior–posterior and medial–lateral axes. The resultant patterning along the neuraxis leads to segmentation of the neuroepithelium into progenitor domains (p0, p1 p2, p3 and pMN) for distinct neuron types in the developing spinal cord. On the basis of several studies it is now believed that this model also applies to macroglial cell specification. Studies carried out by Hochstim and colleagues have demonstrated that three distinct populations of astrocytes arise from the p1, p2 and p3 domains. These subtypes of astrocytes can be identified on the basis of their expression of different transcription factors (PAX6, NKX6.1) and cell surface markers (reelin and SLIT1). The three populations of astrocyte subtypes which have been identified are 1) dorsally located VA1 astrocytes, derived from p1 domain, express PAX6 and reelin 2) ventrally located VA3 astrocytes, derived from p3, express NKX6.1 and SLIT1 and 3) and intermediate white-matter located VA2 astrocyte, derived from the p2 domain, which express PAX6, NKX6.1, reelin and SLIT1. After astrocyte specification has occurred in the developing CNS, it is believed that astrocyte precursors migrate to their final positions within the nervous system before the process of terminal differentiation occurs.
Function
Previously in medical science, the neuronal network was considered the only important function of astrocytes, and they were looked upon as gap fillers. More recently, the function of astrocytes has been reconsidered, and they are now thought to play a number of active roles in the brain, including the secretion or absorption of neural transmitters and maintenance of the blood–brain barrier. Following on this idea the concept of a "tripartite synapse" has been proposed, referring to the tight relationship occurring at synapses among a presynaptic element, a postsynaptic element and a glial element.
Astrocytes are linked by gap junctions, creating an electrically coupled (functional) syncytium. Because of this ability of astrocytes to communicate with their neighbors, changes in the activity of one astrocyte can have repercussions on the activities of others that are quite distant from the original astrocyte.
An influx of Ca2+ ions into astrocytes is the essential change that ultimately generates calcium waves. Because this influx is directly caused by an increase in blood flow to the brain, calcium waves are said to be a kind of hemodynamic response function. An increase in intracellular calcium concentration can propagate outwards through this functional syncytium. Mechanisms of calcium wave propagation include diffusion of calcium ions and IP3 through gap junctions and extracellular ATP signalling. Calcium elevations are the primary known axis of activation in astrocytes, and are necessary and sufficient for some types of astrocytic glutamate release.
Given the importance of calcium signaling in astrocytes, tight regulatory mechanisms for the progression of the spatio-temporal calcium signaling have been developed. Via mathematical analysis it has been shown that localized inflow of Ca2+ ions yields a localized raise in the cytosolic concentration of Ca2+ ions. Moreover, cytosolic Ca2+ accumulation is independent of every intracellular calcium flux and depends on the Ca2+ exchange across the membrane, cytosolic calcium diffusion, geometry of the cell, extracellular calcium perturbation, and initial concentrations.
Bergmann glia
Bergmann glia, a type of glia also known as radial epithelial cells (as named by Camillo Golgi) or Golgi epithelial cells (GCEs; not to be mixed up with Golgi cells), are astrocytes in the cerebellum that have their cell bodies in the Purkinje cell layer and processes that extend into the molecular layer, terminating with bulbous endfeet at the pial surface. Bergmann glia express high densities of glutamate transporters that limit diffusion of the neurotransmitter glutamate during its release from synaptic terminals. Besides their role in early development of the cerebellum, Bergmann glia are also required for the pruning or addition of synapses.
Tripartite synapse
Within the dorsal horn of the spinal cord, activated astrocytes have the ability to respond to almost all neurotransmitters and, upon activation, release a multitude of neuroactive molecules such as glutamate, ATP, nitric oxide (NO), prostaglandins (PG), and D-serine, which in turn influences neuronal excitability. The close association between astrocytes and presynaptic and postsynaptic terminals as well as their ability to integrate synaptic activity and release neuromodulators has been termed the "tripartite synapse". Synaptic modulation by astrocytes takes place because of this 3-part association.
Astrocytomas
Astrocytomas are primary intracranial tumors derived from astrocytes cells of the brain. It is also possible that glial progenitors or neural stem cells give rise to astrocytomas.
Astrocytomas are brain tumors that develop from astrocytes. They may occur in many parts of the brain and sometimes in the spinal cord. They can occur at any age and they primarily occur in males. Astrocytomas are divided into two categories: Low grade (I and II) and High Grade (III and IV). Low grade tumors are more common in children and high grade tumors are more common in adults.
Pilocytic astrocytoma are Grade I tumors. They are considered benign and slow growing tumors. Pilocytic astrocytomas frequently have cystic portions filled with fluid and a nodule, which is the solid portion. Most are located in the cerebellum. Therefore, most symptoms are related to balance or coordination difficulties. They also occur more frequently in children and teens.
Grade II Tumors grow relatively slow but invade surrounding healthy tissue. Usually considered benign but can grow into malignant tumors. Other names that are used are Fibrillary or Protoplasmic astrocytomas. They are prevalent in younger people who are often present with seizures.
Anaplastic astrocytoma is classified as grade III and are malignant tumors. They grow more rapidly than lower grade tumors and tend to invade nearby healthy tissue. Anaplastic astrocytomas recur more frequently than lower grade tumors because of their tendency to spread into surrounding tissue makes them difficult to completely remove surgically.
Glioblastoma Multiforme is also a malignant tumor and classified as a grade IV. Glioblastomas can contain more than one cell type (i.e., astrocytes, oligondroctyes). Also, while one cell type may die off in response to a particular treatment, the other cell types may continue to multiply. Glioblastomas are the most invasive type of glial tumors as they grow rapidly and spread to nearby tissue. Approximately 50% of astrocytomas are glioblastomas and are very difficult to treat.
Neurodevelopment disorders
A relatively new direction has emerged in the field of neurodevelopmental disorders. This view states that astrocyte dysfunction may play a role in the aberration of neuronal circuitry that underlies certain psychiatric disorders, including the autism spectrum disorders (ASDs) and schizophrenia.
Chronic pain
Under normal conditions, pain conduction begins with some noxious signal followed by an action potential carried by nociceptive (pain sensing) afferent neurons, which elicit excitatory postsynaptic potentials (EPSP) in the dorsal horn of the spinal cord. That message is then relayed to the cerebral cortex, where we translate those EPSPs into "pain." Since the discovery of astrocytic influence, our understanding of the conduction of pain has been dramatically complicated. Pain processing is no longer seen as a repetitive relay of signals from body to brain, but as a complex system that can be up- and down-regulated by a number of different factors. One factor at the forefront of recent research is in the pain-potentiating synapse located in the dorsal horn of the spinal cord and the role of astrocytes in encapsulating these synapses. Garrison and co-workers were the first to suggest association when they found a correlation between astrocyte hypertrophy in the dorsal horn of the spinal cord and hypersensitivity to pain after peripheral nerve injury, typically considered an indicator of glial activation after injury. Astrocytes detect neuronal activity and can release chemical transmitters, which in turn control synaptic activity. In the past, hyperalgesia was thought to be modulated by the release of substance P and excitatory amino acids (EAA), such as glutamate, from the presynaptic afferent nerve terminals in the spinal cord dorsal horn. Subsequent activation of AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid), NMDA (N-methyl-D-aspartate) and kainate subtypes of ionotropic glutamate receptors follows. It is the activation of these receptors that potentiates the pain signal up the spinal cord. This idea, although true, is an oversimplification of pain transduction. A litany of other neurotransmitter and neuromodulators, such as calcitonin gene-related peptide (CGRP), adenosine triphosphate (ATP), brain-derived neurotrophic factor (BDNF), somatostatin, vasoactive intestinal peptide (VIP), galanin, and vasopressin are all synthesized and released in response to noxious stimuli. In addition to each of these regulatory factors, several other interactions between pain-transmitting neurons and other neurons in the dorsal horn have added impact on pain pathways.
Two states of persistent pain
After persistent peripheral tissue damage there is a release of several factors from the injured tissue as well as in the spinal dorsal horn. These factors increase the responsiveness of the dorsal horn pain-projection neurons to ensuing stimuli, termed "spinal sensitization," thus amplifying the pain impulse to the brain. Release of glutamate, substance P, and calcitonin gene-related peptide (CGRP) mediates NMDAR activation (originally silent because it is plugged by Mg2+), thus aiding in depolarization of the postsynaptic pain-transmitting neurons (PTN). In addition, activation of IP3 signaling and MAPKs (mitogen-activated protein kinases) such as ERK and JNK, bring about an increase in the synthesis of inflammatory factors that alter glutamate transporter function. ERK also further activates AMPARs and NMDARs in neurons. Nociception is further sensitized by the association of ATP and substance P with their respective receptors (P2X3) and neurokinin 1 receptor (NK1R), as well as activation of metabotropic glutamate receptors and release of BDNF. Persistent presence of glutamate in the synapse eventually results in dysregulation of GLT1 and GLAST, crucial transporters of glutamate into astrocytes. Ongoing excitation can also induce ERK and JNK activation, resulting in release of several inflammatory factors.
As noxious pain is sustained, spinal sensitization creates transcriptional changes in the neurons of the dorsal horn that lead to altered function for extended periods. Mobilization of Ca2+ from internal stores results from persistent synaptic activity and leads to the release of glutamate, ATP, tumor necrosis factor-α (TNF-α), interleukin 1β (IL-1β), IL-6, nitric oxide (NO), and prostaglandin E2 (PGE2). Activated astrocytes are also a source of matrix metalloproteinase 2 (MMP2), which induces pro-IL-1β cleavage and sustains astrocyte activation. In this chronic signaling pathway, p38 is activated as a result of IL-1β signaling, and there is a presence of chemokines that trigger their receptors to become active. In response to nerve damage, heat shock proteins (HSP) are released and can bind to their respective TLRs, leading to further activation.
Other pathologies
Other clinically significant pathologies involving astrocytes include astrogliosis and astrocytopathy.
Recent studies
A recent study, done in November 2010 and published March 2011, was done by a team of scientists from the University of Rochester and University of Colorado School of Medicine. They did an experiment to attempt to repair trauma to the Central Nervous System of an adult rat by replacing the glial cells. When the glial cells were injected into the injury of the adult rat’s spinal cord, astrocytes were generated by exposing human glial precursor cells to bone morphogenetic protein (Bone morphogenetic protein is important because it is considered to create tissue architecture throughout the body). So, with the bone protein and human glial cells combined, they promoted significant recovery of conscious foot placement, axonal growth, and obvious increases in neuronal survival in the spinal cord laminae. On the other hand, human glial precursor cells and astrocytes generated from these cells by being in contact with ciliary neurotrophic factors, failed to promote neuronal survival and support of axonal growth at the spot of the injury.
One study done in Shanghai had two types of hippocampal neuronal cultures: In one culture, the neuron was grown from a layer of astrocytes and the other culture was not in contact with any astrocytes, but they were instead fed a glial conditioned medium (GCM), which inhibits the rapid growth of cultured astrocytes in the brains of rats in most cases. In their results they were able to see that astrocytes had a direct role in Long-term potentiation with the mixed culture (which is the culture that was grown from a layer of astrocytes) but not in GCM cultures.
Recent studies have shown that astrocytes play an important function in the regulation of neural stem cells. Research from the Schepens Eye Research Institute at Harvard shows the human brain to abound in neural stem cells, which are kept in a dormant state by chemical signals (ephrin-A2 and ephrin-A3) from the astrocytes. The astrocytes are able to activate the stem cells to transform into working neurons by dampening the release of ephrin-A2 and ephrin-A3.
In a study published in a 2011 issue of Nature Biotechnology a group of researchers from the University of Wisconsin reports that it has been able to direct embryonic and induced human stem cells to become astrocytes.
A 2012 study of the effects of marijuana on short term memories found that THC activates CB1 receptors of astrocytes which cause receptors for AMPA to be removed from the membranes of associated neurons.
Classification
There are several different ways to classify astrocytes.
Lineage and antigenic phenotype
These have been established by classic work by Raff et al. in early 1980s on Rat optic nerves.