
Scientific name Arachnida | Phylum Arthropoda | |
 | ||
Lower classifications | ||
Arachnid diversity eight legged diversity spiders and their kin
Arachnids are a class (Arachnida) of joint-legged invertebrate animals (arthropods), in the subphylum Chelicerata. All arachnids have eight legs, although the front pair of legs in some species has converted to a sensory function, while in other species, different appendages can grow large enough to take on the appearance of extra pairs of legs. The term is derived from the Greek word ἀράχνη (aráchnē), meaning "spider". Spiders are the largest order in the class.
Contents
- Arachnid diversity eight legged diversity spiders and their kin
- 9 amazing new arachnid species
- Morphology
- Locomotion
- Physiology
- Diet and digestive system
- Senses
- Reproduction
- Phylogeny
- Subdivisions
- Acari
- Amblypygi
- Araneae
- Haptopoda
- Opiliones
- Palpigradi
- Phalangiotarbi
- Pseudoscorpions
- Ricinulei
- Schizomida
- Scorpiones
- Solifugae
- Thelyphonida
- Trigonotarbida
- Uraraneida
- References

Almost all extant arachnids are terrestrial, living mainly on land. However, some inhabit freshwater environments and, with the exception of the pelagic zone, marine environments as well. They comprise over 100,000 named species, including spiders, Scorpions, harvestmen, ticks, mites, and solifuges.
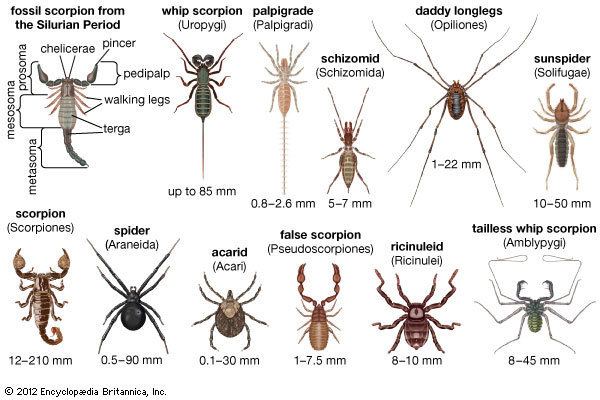
9 amazing new arachnid species
Morphology

Almost all adult arachnids have eight legs, and arachnids may be easily distinguished from insects by this fact, since insects have six legs. However, arachnids also have two further pairs of appendages that have become adapted for feeding, defense, and sensory perception. The first pair, the chelicerae, serve in feeding and defense. The next pair of appendages, the pedipalps, have been adapted for feeding, locomotion, and/or reproductive functions. In Solifugae, the palps are quite leg-like, so that these animals appear to have ten legs. The larvae of mites and Ricinulei have only six legs; a fourth pair usually appears when they moult into nymphs. However, mites are variable: as well as eight, there are adult mites with six or even four legs.

Arachnids are further distinguished from insects by the fact they do not have antennae or wings. Their body is organized into two tagmata, called the prosoma, or cephalothorax, and the opisthosoma, or abdomen. The cephalothorax is derived from the fusion of the cephalon (head) and the thorax, and is usually covered by a single, unsegmented carapace. The abdomen is segmented in the more primitive forms, but varying degrees of fusion between the segments occur in many groups. It is typically divided into a preabdomen and postabdomen, although this is only clearly visible in scorpions, and in some orders, such as the Acari, the abdominal sections are completely fused.

Like all arthropods, arachnids have an exoskeleton, and they also have an internal structure of cartilage-like tissue, called the endosternite, to which certain muscle groups are attached. The endosternite is even calcified in some Opiliones.
Locomotion
Most arachnids lack extensor muscles in the distal joints of their appendages. Spiders and whipscorpions extend their limbs hydraulically using the pressure of their hemolymph. Solifuges and some harvestmen extend their knees by the use of highly elastic thickenings in the joint cuticle. Scorpions, pseudoscorpions and some harvestmen have evolved muscles that extend two leg joints (the femur-patella and patella-tibia joints) at once. The equivalent joints of the pedipalps of scorpions though, are extended by elastic recoil.
Physiology
There are characteristics that are particularly important for the terrestrial lifestyle of arachnids, such as internal respiratory surfaces in the form of tracheae, or modification of the book gill into a book lung, an internal series of vascular lamellae used for gas exchange with the air. While the tracheae are often individual systems of tubes, similar to those in insects, ricinuleids, pseudoscorpions, and some spiders possess sieve tracheae, in which several tubes arise in a bundle from a small chamber connected to the spiracle. This type of tracheal system has almost certainly evolved from the book lungs, and indicates that the tracheae of arachnids are not homologous with those of insects.
Further adaptations to terrestrial life are appendages modified for more efficient locomotion on land, internal fertilisation, special sensory organs, and water conservation enhanced by efficient excretory structures as well as a waxy layer covering the cuticle.
The excretory glands of arachnids include up to four pairs of coxal glands along the side of the prosoma, and one or two pairs of Malpighian tubules, emptying into the gut. Many arachnids have only one or the other type of excretory gland, although several do have both. The primary nitrogenous waste product in arachnids is guanine.
Arachnid blood is variable in composition, depending on the mode of respiration. Arachnids with an efficient tracheal system do not need to transport oxygen in the blood, and may have a reduced circulatory system. In scorpions and some spiders, however, the blood contains haemocyanin, a copper-based pigment with a similar function to haemoglobin in vertebrates. The heart is located in the forward part of the abdomen, and may or may not be segmented. Some mites have no heart at all.
Diet and digestive system
Arachnids are mostly carnivorous, feeding on the pre-digested bodies of insects and other small animals. Only in the harvestmen and among mites, such as the house dust mite, is there ingestion of solid food particles, and thus exposure to internal parasites, although it is not unusual for spiders to eat their own silk. Several groups secrete venom from specialized glands to kill prey or enemies. Several mites and ticks are parasites, some of which are carriers of disease.
Arachnids produce digestive juices in their stomachs, and use their pedipalps and chelicerae to pour them over their dead prey. The digestive juices rapidly turn the prey into a broth of nutrients, which the arachnid sucks into a pre-buccal cavity located immediately in front of the mouth. Behind the mouth is a muscular, sclerotised pharynx, which acts as a pump, sucking the food through the mouth and on into the oesophagus and stomach. In some arachnids, the oesophagus also acts as an additional pump.
The stomach is tubular in shape, with multiple diverticula extending throughout the body. The stomach and its diverticula both produce digestive enzymes and absorb nutrients from the food. It extends through most of the body, and connects to a short sclerotised intestine and anus in the hind part of the abdomen.
Senses
Arachnids have two kinds of eyes: the lateral and median ocelli. The lateral ocelli evolved from compound eyes and may have a tapetum, which enhances the ability to collect light. With the exception of scorpions, which can have up to five pairs of lateral ocelli, there are never more than three pairs present. The median ocelli develop from a transverse fold of the ectoderm. The ancestors of modern arachnids probably had both types, but modern ones often lack one type or the other. The cornea of the eye also acts as a lens, and is continuous with the cuticle of the body. Beneath this is a transparent vitreous body, and then the retina and, if present, the tapetum. In most arachnids, the retina probably does not have enough light sensitive cells to allow the eyes to form a proper image.
In addition to the eyes, almost all arachnids have two other types of sensory organs. The most important to most arachnids are the fine sensory hairs that cover the body and give the animal its sense of touch. These can be relatively simple, but many arachnids also possess more complex structures, called trichobothria.
Finally, slit sense organs are slit-like pits covered with a thin membrane. Inside the pit, a small hair touches the underside of the membrane, and detects its motion. Slit sense organs are believed to be involved in proprioception, and possibly also hearing.
Reproduction
Arachnids may have one or two gonads, which are located in the abdomen. The genital opening is usually located on the underside of the second abdominal segment. In most species, the male transfers sperm to the female in a package, or spermatophore. Complex courtship rituals have evolved in many arachnids to ensure the safe delivery of the sperm to the female.
Arachnids usually lay yolky eggs, which hatch into immatures that resemble adults. Scorpions, however, are either ovoviviparous or viviparous, depending on species, and bear live young. In most arachnids only the females provide parental care, with harvestmen being one of the few exceptions.
Phylogeny
The phylogenetic relationships among the main subdivisions of arthropods have been the subject of considerable research and dispute for many years. A consensus emerged from about 2010 onwards, based on both morphological and molecular evidence. Extant (living) arthropods are a monophyletic group and are divided into three main clades: chelicerates (including arachnids), pancrustaceans (the paraphyletic crustaceans plus insects and their allies), and myriapods (centipedes, millipedes and allies). The three groups are related as shown in the cladogram below. Including fossil taxa does not fundamentally alter this view, although it introduces some additional basal groups.
The extant chelicerates comprise two marine groups: sea spiders and horseshoe crabs, and the terrestrial arachnids. These are related as shown below. (Pycnogonida (sea spiders) may be excluded from the chelicerates, which are then identified as the group labelled "Euchelicerata".)
Discovering relationships within the arachnids has proved difficult as of March 2016, with successive studies producing different results. A study in 2014, based on the largest set of molecular data to date, concluded that there were systematic conflicts in the phylogenetic information, particularly affecting the orders Acariformes, Parasitiformes and Pseudoscorpiones, which have had much faster evolutionary rates. Analyses of the data using sets of genes with different evolutionary rates produced mutually incompatible phylogenetic trees. The authors favoured relationships shown by more slowly evolving genes, which demonstrated the monophyly of Chelicerata, Euchelicerata and Arachnida, as well as of some clades within the arachnids. The diagram below summarizes their conclusions, based largely on the 200 most slowly evolving genes; dashed lines represent uncertain placements.
Tetrapulmonata, here consisting of Araneae, Amblypygi and Thelyphonida (Schizomida was not included in the study), received strong support. The addition of Scorpiones to produce a clade called Arachnopulmonata was also well supported. Pseudoscorpiones may also belong here, possibly as the sister of Scorpiones. Somewhat unexpectedly, there was support for a clade comprising Opiliones, Ricinulei and Solifugae, a combination not found in most other studies.
Subdivisions
The subdivisions of the arachnids are usually treated as orders. Historically, mites and ticks were treated as a single order, Acari. However, molecular phylogenetic studies suggest that the two groups do not form a single clade, with morphological similarities being due to convergence. They are now usually treated as two separate taxa – Acariformes, mites, and Parasitiformes, ticks – which may be ranked as orders or superorders. The arachnid subdivisions are listed below alphabetically; numbers of species are approximate.
It is estimated that 98,000 arachnid species have been described, and that there may be up to 600,000 in total.
Acari
Acari or Acarina is a taxon of arachnids that contains mites and ticks. Its fossil history goes back to the Devonian period, although there is also a questionable Ordovician record. The Devonian period was the time frame in which certain species of animals developed legs. In most modern treatments, the Acari is considered a subclass of Arachnida and is composed of two or three orders or superorders: Acariformes, Parasitiformes, and Opilioacariformes. Most acarines are minute to small (e.g. 0.080–1.00 mm), but the giants of the Acari (some ticks and red velvet mites) may reach lengths of 10–20 mm. It is estimated that over 50,000 species have been described (as of 1999) and that a million or more species are currently living. The study of mites and ticks is called acarology.
Only the faintest traces of primary segmentation remain in mites, the prosoma and opisthosoma being insensibly fused, and a region of flexible cuticle (the cirumcapitular furrow) separates the chelicerae and pedipalps from the rest of the body. This anterior body region is called the gnathosoma (or capitulum) and is also found in the Ricinulei. The remainder of the body is called the idiosoma and is unique to mites. Most adult mites have four pairs of legs, like other arachnids, but some have fewer. For example, gall mites like Phyllocoptes variabilis (superfamily Eriophyioidea) have a wormlike body with only two pairs of legs; some parasitic mites have only one or three pairs of legs in the adult stage. Larval and prelarval stages have a maximum of three pairs of legs; adult mites with only three pairs of legs may be called 'larviform'.
Acarine ontogeny consists of an egg, a prelarval stage (often absent), a larval stage (hexapod except in Eriophyoidea, which have only two pairs of legs), and a series of nymphal stages. Larvae (and prelarvae) have a maximum of three pairs of legs (legs are often reduced to stubs or absent in prelarvae); the fourth pair of legs is added at the first nymphal stage.
Acarines live in practically every habitat, and include aquatic (freshwater and sea water) and terrestrial species. They outnumber other arthropods in the soil organic matter and detritus. Many are parasitic, and they affect both vertebrates and invertebrates. Most parasitic forms are external parasites, while the free living forms are generally predaceous and may even be used to control undesirable arthropods. Others are detritivores that help to break down forest litter and dead organic matter such as skin cells. Others still are plant feeders and may damage crops. Damage to crops is perhaps the most costly economic effect of mites, especially by the spider mites and their relatives (Tetranychoidea), earth mites (Penthaleidae), thread-footed mites (Tarsonemidae) and the gall and rust mites (Eriophyoidea). Some parasitic forms affect humans and other mammals, causing damage by their feeding, and can even be vectors of diseases such as scrub typhus and rickettsial pox. A well-known effect of mites on humans is their role as an allergen and the stimulation of asthma in people affected by the respiratory disease. The use of predatory mites (e.g. Phytoseiidae) in pest control and herbivorous mites that attack weeds is also important. An unquantified, but major positive contribution of the Acari is their normal functioning in ecosystems, especially their roles in the decomposer subsystem.
Amblypygi
Amblypygids are also known as tailless whip scorpions or cave spiders. Approximately 5 families, 17 genera and 136 species have been described. They are found in tropical and subtropical regions worldwide. Some species are subterranean; many are nocturnal. During the day, they may hide under logs, bark, stones, or leaves. They prefer a humid environment. Amblypygids may range from 5 to 40 mm. Their bodies are broad and highly flattened and the first pair of legs (the first walking legs in most arachnid orders) are modified to act as sensory organs. (Compare solifugids, uropygids, and schizomids.) These very thin modified legs can extend several times the length of body. They have no silk glands or venomous fangs, but can have prominent pincer-like pedipalps. Amblypygids often move about sideways on their six walking legs, with one "whip" pointed in the direction of travel while the other probes on either side of them. Prey are located with these "whips", captured with pedipalps, then torn to pieces with chelicerae. Fossilised amblypygids have been found dating back to the Carboniferous period.
Amblypygids, particularly the species Phrynus marginemaculatus and Damon diadema, are thought to be one of the few species of arachnids that show signs of social behavior. Research conducted at Cornell University by entomologists suggests that a mother amblypygid comforts her young by gently caressing the offspring with her feelers. Further, when two or more siblings were placed in an unfamiliar environment, such as a cage, they would seek each other out and gather back in a group.
Araneae
Araneae, or spiders, are the most familiar of the arachnids, and the most species-rich with around 40,000 described species. All spiders produce silk, a thin, strong protein strand extruded by the spider from spinnerets most commonly found on the end of the abdomen. Many species use it to trap insects in webs, although there are many species that hunt freely. Silk can be used to aid in climbing, form smooth walls for burrows, build egg sacs, wrap prey, temporarily hold sperm, and even fly, among other applications.
All spiders except those in the families Uloboridae and Holarchaeidae, and in the suborder Mesothelae (together about 350 species) can inject venom to protect themselves or to kill and liquefy prey. Only about 200 species, however, have bites that can pose health problems to humans. Many larger species' bites may be painful, but will not produce lasting health concerns.
Spiders are found all over the world, from the tropics to the Arctic, with some extreme species even living underwater in silken domes that they supply with air, and on the tops of the highest mountains.
Haptopoda
Haptopoda is an extinct order known exclusively from a few specimens from the Upper Carboniferous of the United Kingdom. It is monotypic, i.e. has only one species, Plesiosiro madeleyi. Relationships with other arachnids are obscure, but closest relatives may be the Amblypygi, Thelyphonida and Schizomida of the tetrapulmonate clade – a result which has been reflected in cladistic analyses.
Opiliones
Opiliones (formerly Phalangida, and better known as harvestmen or daddy longlegs) are arachnids that are harmless to people and are known for their exceptionally long walking legs, compared to their body size. As of December 2011, over 6,500 species of harvestmen have been discovered worldwide. The order Opiliones is divided into five suborders: Cyphophthalmi, Eupnoi, Dyspnoi, Laniatores, and the recently described Tetrophthalmi. Well-preserved fossils have been found in the 410-million year old Rhynie cherts of Scotland and 305-million-year-old rocks from France; they look surprisingly modern, suggesting that the basic structure of the harvestmen has not changed much since then.
The difference between harvestmen and spiders is that in harvestmen the two main body sections (the abdomen or opisthosoma with ten segments and the cephalothorax or prosoma) are nearly joined, so that they appear to be one oval structure. In more advanced species, the first five abdominal segments are often fused into a dorsal shield called the scutum, which is normally fused with the carapace. Sometimes this shield is only present in males. The two hindmost abdominal segments may be reduced or separated in the middle on the surface to form two plates lying next to each other. The second pair of legs is longer than the others and works as antennae. They have a single pair of eyes in the middle of their heads, oriented sideways. They have a pair of prosomatic scent glands that secrete a peculiar smelling fluid when disturbed. Harvestmen do not have spinnerets and do not possess poison glands, posing absolutely no danger to humans. They breathe through tracheae. Between the base of the fourth pair of legs and the abdomen is a pair of spiracles, one opening on each side. In more active species, spiracles are also found upon the tibia of the legs. They have a gonopore on the ventral cephalothorax, and copulation is direct, as the male has a penis (while the female has an ovipositor).
Typical body length does not exceed 7 millimetres (0.28 in) even in the largest species. However, leg span is much larger and can exceed 160 mm (6.3 in). Most species live for a year. Many species are omnivorous, eating primarily small insects and all kinds of plant material and fungi; some are scavengers of the decays of any dead animal, bird dung and other fecal material. They are mostly nocturnal and coloured in hues of brown, although there are a number of diurnal species that have vivid patterns in yellow, green and black with varied reddish and blackish mottling and reticulation.
Palpigradi
Palpigradi, commonly known as "microwhip scorpions", are tiny cousins of the uropygid, or whip scorpion, no more than 3 mm in length. They have a thin, pale, segmented carapace that terminates in a whip-like flagellum, made up of 15 segments. The carapace is divided into two plates between the third and fourth leg set. They have no eyes. Some species have three pairs of book lungs, while others have no respiratory organs at all. Approximately 80 species of Palpigradi have been described worldwide, in the families Eukoeneniidae and Prokoeneniidae, with a total of seven genera.
They are believed to be predators like their larger relatives, feeding on minuscule insects in their habitat. Their mating habits are unknown, except that they lay only a few relatively large eggs at a time. Microwhip scorpions need a damp environment to survive, and they always hide from light, so they are commonly found in the moist earth under buried stones and rocks. They can be found on every continent, except in Arctic and Antarctic regions.
Phalangiotarbi
Phalangiotarbi is an extinct arachnid order known exclusively from the Palaeozoic (Devonian to Permian) of Europe and North America. The affinities of phalangiotarbids are obscure, with most authors favouring affinities with Opiliones (harvestmen) and/or Acari (mites and ticks). Phalangiotarbi has been recently proposed to be sister group to (Palpigradi+Tetrapulmonata): the taxon Megoperculata sensu Shultz (1990).
Pseudoscorpions
Pseudoscorpions are small arthropods with a flat, pear-shaped body and pincers that resemble those of scorpions. They range from 2 to 8 mm (0.079 to 0.315 in) long. The opisthosoma is made up of twelve segments, each guarded by plate-like tergites above and sternites below. The abdomen is short and rounded at the rear, rather than extending into a segmented tail and stinger like true scorpions. The colour of the body can be yellowish-tan to dark-brown, with the paired claws often a contrasting colour. They may have two, four or no eyes. They have two very long pedipalps with palpal chelae (pincers) that strongly resemble the pincers found on a scorpion. The pedipalps generally consist of an immobile "hand" and "finger", with a separate movable finger controlled by an adductor muscle. A venom gland and duct are usually located in the mobile finger; the poison is used to capture and immobilise the pseudoscorpion'S prey. During digestion, pseudoscorpions pour a mildly corrosive fluid over the prey, then ingest the liquefied remains. Pseudoscorpions spin silk from a gland in their jaws to make disk-shaped cocoons for mating, molting, or waiting out cold weather. Another trait they share with their closest relatives, the spiders, is breathing through spiracles. Most spiders have one pair of spiracles, and one of book lungs, but pseudoscorpions do not have book lungs.
There are more than 2,000 species of pseudoscorpions recorded. They range worldwide, even in temperate to cold regions, but have their most dense and diverse populations in the tropics and subtropics. The fossil record of pseudoscorpions dates back over 380 million years, to the Devonian period, near the time when the first land-animal fossils appear.
During the elaborate mating dance, the male of some pseudoscorpion species pulls a female over a spermatophore previously laid upon a surface. In other species, the male also pushes the sperm into the female genitals using the forelegs. The female carries the fertilised eggs in a brood pouch attached to her abdomen, and the young ride on the mother for a short time after they hatch. Up to two dozen young are hatched in a single brood; there may be more than one brood per year. The young go through three molts over the course of several years before reaching adulthood. Adult pseudoscorpions live 2 to 3 years. They are active in the warm months of the year, overwintering in silken cocoons when the weather grows cold.
Pseudoscorpions are generally beneficial to humans since they prey on clothes moth larvae, carpet beetle larvae, booklice, ants, mites, and small flies. They are small and inoffensive, and are rarely seen due to their size. They usually enter the home by "riding along" with larger insects (known as phoresy), or are brought in with firewood. They are often observed in bathrooms or laundry rooms, since they seek humidity. They may sometimes be found feeding on mites under the wing covers of certain beetles.
Ricinulei
Ricinulei (hooded tickspiders) are 5–10 mm long. Their most notable feature is a "hood" that can be raised and lowered over the head; when lowered, it covers the mouth and the chelicerae. Ricinulei have no eyes. The pedipalps end in pincers that are small relative to their bodies, when compared to those of the related orders of scorpions and pseudoscorpions. The heavy-bodied abdomen forms a narrow pedicel, or waist, where it attaches to the prosoma. In males, the third pair of legs are modified to form copulatory organs. Malpighian tubules and a pair of coxal glands make up the excretory system. They have no lungs, as gas exchange takes place through the trachea.
Ricinulei are predators, feeding on other small arthropods. Little is known about their mating habits; the males have been observed using their modified third leg to transfer a spermatophore to the female. The eggs are carried under the mother's hood, until the young hatch into six-legged "larva", which later molt into their adult forms. Ricinulei require moisture to survive. Approximately 57 species of ricinuleids have been described worldwide, all in a single family that contains three genera.
Schizomida
Schizomida is an order of arachnids that tend to live in the top layer of soils. Schizomids present the prosoma covered by a large protopeltidium and smaller, paired, mesopeltidia and metapeltidia. There are no eyes. The opisthosoma is a smooth oval of 12 recognisable somites. The first is reduced and forms the pedicel. The last three are much constricted, forming the pygidium. The last somite bears the flagellum, which in this order is short and consists of not more than four segments.
The name means "split or cleaved middle", referring to the way the cephalothorax is divided into two separate plates. Like the Thelyphonida, Amblypygi, and Solifugae, the schizomids use only six legs for walking, having modified their first two legs to serve as sensory organs. They also have large well-developed pedipalps (pincers) just behind the sensory legs.
Scorpiones
Scorpions (formally the order Scorpiones) are characterised by a metasoma (tail) comprising six segments, the last containing the scorpion's anus and bearing the telson (the sting). The telson, in turn, consists of the vesicle, which holds a pair of venom glands and the hypodermic aculeus, the venom-injecting barb. The abdomen's front half, the mesosoma, is made up of six segments. The first segment contains the sexual organs as well as a pair of vestigial and modified appendages forming a structure called the genital operculum. The second segment bears a pair of featherlike sensory organs known as the "pectines"; the final four segments each contain a pair of book lungs. The mesosoma is armored with chitinous plates, known as tergites on the upper surface and sternites on the lower surface.
The cuticle of scorpions is covered with hairs in some places that act like balance organs. An outer layer that makes them fluorescent green under ultraviolet light is called the hyaline layer. Newly molted scorpions do not glow until after their cuticle has hardened. The fluorescent hyaline layer can be intact in fossil rocks that are hundreds of millions of years old.
Scorpions are opportunistic predators of small arthropods and insects. They use their chelae (pincers) to catch the prey initially. Depending on the toxicity of their venom and size of their claws, they will then either crush the prey or inject it with neurotoxic venom. The neurotoxins consist of a variety of small proteins as well as sodium and potassium cations, which serve to interfere with neurotransmission in the victim. Scorpions use their venom to kill or paralyze their prey so that it can be eaten; in general, it is fast acting, allowing for effective prey capture. Scorpion venoms are optimised for action on other arthropods and therefore most scorpions are relatively harmless to humans; stings produce only local effects (such as pain, numbness or swelling). A few scorpion species, however, mostly in the family Buthidae, can be dangerous to humans. The scorpion that is responsible for the most human deaths is Androctonus australis, or fat-tailed scorpion of North Africa. The toxicity of A. australis's venom is roughly half that of the deathstalker (Leiurus quinquestriatus), but since A. australis injects quite a bit more venom into its prey, it is the most deadly to humans. Human deaths normally occur in the young, elderly, or infirm; scorpions are generally unable to deliver enough venom to kill healthy adults. Some people, however, may be allergic to the venom of some species, in which case the scorpion's sting can more likely kill. A primary symptom of a scorpion sting is numbing at the injection site, sometimes lasting for several days. It has been found that scorpions have two types of venom: a translucent, weaker venom designed to stun only, and an opaque, more potent venom designed to kill heavier threats.
Unlike the majority of Arachnida species, scorpions are viviparous. The young are born one by one, and the brood is carried about on its mother's back until the young have undergone at least one moult. The young generally resemble their parents, requiring between five and seven moults to reach maturity. Scorpions have quite variable lifespans and the lifespan of most species is not known. The age range appears to be approximately 4–25 years (25 years being the maximum reported life span in the giant desert hairy scorpion, Hadrurus arizonensis). They are nocturnal and fossorial, finding shelter during the day in the relative cool of underground holes or undersides of rocks and coming out at night to hunt and feed. Scorpions prefer to live in areas where the temperature is 20–37 °C (68–99 °F), but may survive in the temperature range of 14–45 °C (57–113 °F).
Scorpions have been found in many fossil records, including coal deposits from the Carboniferous Period and in marine Silurian deposits. They are thought to have existed in some form since about 450 to 425 million years ago. They are believed to have an oceanic origin, with gills and a claw-like appendage that enabled them to hold onto rocky shores or seaweed.
Solifugae
Solifugae is a group of 900 species of arachnids, commonly known as camel spiders, wind scorpions, and sun spiders. The name derives from Latin, and means those that flee from the sun. Most Solifugae live in tropical or semitropical regions where they inhabit warm and arid habitats, but some species have been known to live in grassland or forest habitats. The most distinctive feature of Solifugae is their large chelicerae. Each of the two chelicerae are composed of two articles forming a powerful pincer; each article bears a variable number of teeth. Males in all families but Eremobatidae possess a flagellum on the basal article of the chelicera. Solifugae also have long pedipalps, which function as sense organs similar to insects' antennae and give the appearance of the two extra legs. Pedipalps terminate in reversible adhesive organs.
Solifugae are carnivorous or omnivorous, with most species feeding on termites, darkling beetles, and other small arthropods; however, solifugae have been videotaped consuming larger prey, such as lizards. Prey is located with the pedipalps and killed and cut into pieces by the chelicerae. The prey is then liquefied and the liquid ingested through the pharynx. Reproduction can involve direct or indirect sperm transfer; when indirect, the male emits a spermatophore on the ground and then inserts it with his chelicerae in the female's genital pore.
Thelyphonida
The Thelyphonida (formerly Uropygi), commonly known as vinegarroons or whip scorpions, range from 25 to 85 mm in length; the largest species, of the genus Mastigoproctus, reaches 85 mm (3.3 in). Like the related orders Schizomida, Amblypygi, and Solifugae, the vinegarroons use only six legs for walking, having modified their first two legs to serve as antennae-like sensory organs. Many species also have very large scorpion-like pedipalps (pincers). They have one pair of eyes at the front of the cephalothorax and three on each side of the head. Whip scorpions have no poison glands, but they do have glands near the rear of their abdomen that can spray a combination of acetic acid and octanoic acid when they are bothered. Other species spray formic acid or chlorine. As of 2006, over 100 species have been described worldwide.
Whip scorpions are carnivorous, nocturnal hunters feeding mostly on insects but sometimes on worms and slugs. The prey is crushed between special teeth on the inside of the trochanters (the second segment of the leg) of the front legs. They are valuable in controlling cockroach and cricket populations.
Males secrete a sperm sac, which is transferred to the female. Up to 35 eggs are laid in a burrow, within a mucous membrane that preserves moisture. Mothers stay with the eggs and do not eat. The white young that hatch from the eggs climb onto their mother's back and attach themselves there with special suckers. After the first molt, they look like miniature whip scorpions, and leave the burrow; the mother dies soon after. The young grow slowly, going through three molts in about three years before reaching adulthood.
Vinegarroons are found in tropical and subtropical areas worldwide, usually in underground burrows that they dig with their pedipalps. They may also burrow under logs, rotting wood, rocks, and other natural debris. They enjoy humid, dark places and avoid the light.
Trigonotarbida
The order Trigonotarbida is an extinct group of arachnids whose fossil record extends from the Silurian to the Lower Permian. They are known from several localities in North Asia, North America and Argentina. They superficially resemble spiders, to which they were clearly related - most cladistic analyses recover them in a clade with Thelyphonida, Schizomida, Amblypygi and Araneae.
These early arachnids seem to have been adapted to stalking prey on the ground. They have been found within the very structure of ground-dwelling plants, possibly where they hid to await their prey. Trigonotarbids are currently among the oldest known land arthropods. They lack silk glands on the opisthosoma and cheliceral poison glands, and most likely represented independent offshoots of the Arachnida.
Uraraneida
Uraraneida is a small extinct order of arachnids consisting of two fossil species found in the Devonian and the Permian. They were at one time identified as spiders, and are clearly closely related to them. However, they differ in a several respects. Silk is produced from spigots borne along the rear edges of plates on the underside of the abdomen, not on appendage-like spinnerets, as in spiders. They also have a long, jointed "tail" or flagellum at the end of the abdomen, after the anus, a feature lacking in spiders but present in some other arachnids, such as whip scorpions.