
Days 16 FMA 69070 | MeSH A16.254.425.273 | |
 | ||
Ectoderm is one of the three primary germ layers in the very early embryo. The other two layers are the mesoderm (middle layer) and endoderm (most proximal layer), with the ectoderm as the most exterior (or distal) layer. It emerges and originates from the outer layer of germ cells. The word ectoderm comes from the Greek ektos meaning "outside", and derma, meaning "skin."
Contents
- History
- Initial appearance
- Early development
- Gastrulation
- Later development
- Neurulation
- Organogenesis
- Ectodermal dysplasia
- References
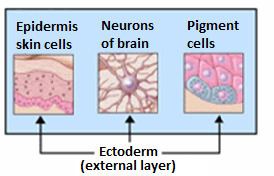
Generally speaking, the ectoderm differentiates to form the nervous system (spine, peripheral nerves and brain), tooth enamel and the epidermis (the outer part of integument). It also forms the lining of mouth, anus, nostrils, sweat glands, hair and nails.
In vertebrates, the ectoderm has three parts: external ectoderm (also known as surface ectoderm), the neural crest, and neural tube. The latter two are known as neuroectoderm.
History
Heinz Christian Pander, a Baltic German–Russian biologist, has been credited for the discovery of the three germ layers that form during embryogenesis. Pander received his doctorate in zoology from the University of Wurzburg in 1817. He began his studies in embryology using chicken eggs, which allowed for his discovery of the ectoderm, mesoderm and endoderm. Due to his findings, Pander is sometimes referred to as the "founder of embryology".
Pander's work of the early embryo was continued by a Prussian–Estonian biologist named Karl Ernst von Baer. Baer took Pander's concept of the germ layers and through extensive research of many different types of species, he was able to extend this principle to all vertebrates. Baer also received credit for the discovery of the blastula. Baer published his findings, including his germ layer theory, in a textbook which translates to On the Development of Animals which he released in 1828.
Initial appearance
The ectoderm can first be observed in amphibians and fish during the later stages of a process called gastrulation. At the start of this process, the developing embryo has divided into many cells separating the embryo, which is now a hollow sphere of cells called the blastula, into two parts, the animal hemisphere and vegetal hemisphere. It is the animal hemisphere of the blastula that will eventually become the ectoderm.
Early development
Like the other two germ layers, mesoderm and endoderm, the ectoderm forms shortly after the egg is fertilized, and rapid cell division initiates. The epidermis of the skin originates from the less dorsal ectoderm which surrounds the neuroectoderm at the early gastrula stage of embryonic development. The position of the ectoderm relative to the other germ layers of the embryo is governed by "selective affinity", meaning that the inner surface of the ectoderm has a strong (positive) affinity for the mesoderm, and a weak (negative) affinity for the endoderm layer. This selective affinity changes during different stages of development. The strength of the attraction between two surfaces of two germ layers is determined by the amount and type of cadherin molecules present on the cells' surface. For example, the expression of N-cadherin is crucial to maintaining separation of precursor neural cells from precursor epithelial cells. The ectoderm is instructed to become the nervous system by the notochord, which is typically positioned above it.
Gastrulation
During the process of gastrulation, a special type of cells called bottle cells invaginates a hole on the surface of the blastula which is called the dorsal lip of the blastopore. Once this lip has been established, the bottle cells will extend inward and migrate along the inner wall of the blastula known as the roof of the blastocoel. The once superficial cells of the animal pole are destined to become the cells of the middle germ layer called the mesoderm. Through the process of radial extension, cells of the animal pole that were once several layers thick divide to from a thin layer. At the same time, when this thin layer of dividing cells reaches the dorsal lip of the blastopore, another process occurs termed convergent extension. During convergent extension, cells that approach the lip intercalate mediolaterally, in such a way that cells are pulled over the lip and inside the embryo. These two processes allow for the prospective mesoderm cells to be placed between the ectoderm and the endoderm. Once convergent extension and radial intercalation are underway, the rest of the vegetal pole, which will become endoderm cells, is completely engulfed by the prospective ectoderm, as these top cells undergo epiboly, where the ectoderm cells divide in a way to form one layer. This creates a uniform embryo composed of the three germ layers in their respective positions.
Later development
Once there is an embryo with three established germ layers, differentiation among these three layers proceeds. The next event that will take place within the ectoderm is the process of neurulation, which results in the formation of the neural tube, neural crest cells and the epidermis. It is these three components of the ectoderm that will each give rise to a particular set of cells. The neural tube cells will become the central nervous system, neural crest cells will become the peripheral and enteric nervous system, along with melanocytes, facial cartilage and the dentin of teeth, and the epidermal cell region will give rise to epidermis, hair, nails, sebaceous glands, olfactory and mouth epithelium, as well as eyes.
Neurulation
Neurulation proceeds by primary and secondary neurulation, both positioning neural crest cells between a superficial epidermal layer and a deep neural tube. During primary neurulation, the notochord cells of the mesoderm signal the adjacent, superficial ectoderm cells to reposition themselves in a columnar pattern to form cells of the ectodermal neural plate. As the cells continue to elongate, a group of cells immediately above the notochord change their shape, forming a wedge in the ectodermal region. These special cells are called medial hinge cells (MHP). Now, as the ectoderm continues to elongate, the ectodermal cells of the neural plate fold inward. The inward folding of the ectoderm by virtue of mainly cell division continues until another group of cells form within the neural plate. These cells are termed dorsolateral hinge cells (DLHP), and once formed, the inward folding of the ectoderm stops. The DLHP cells function in a similar fashion as MHP cells regarding their wedge like shape, however, the DLHP cells result in the ectoderm converging. This convergence is led by ectodermal cells above the DLHP cells known as the neural crest. The neural crest cells eventually pull the adjacent ectodermal cells together, which leaves neural crest cells between the prospective epidermis and hollow, neural tube.
Organogenesis
All of the organs that rise from the ectoderm such as the nervous system, teeth, hair and many exocrine glands, originate from two adjacent tissue layers: the epithelium and the mesenchyme. Several signals mediate the organogenesis of the ectoderm such as: FGF, TGFβ, Wnt, and regulators from the hedgehog family. The specific timing and manner that the ectodermal organs form is dependent on the invagination of the epithelial cells. FGF-9 is an important factor during the initiation of tooth germ development. The rate of epithelial invagination in significantly increased by action of FGF-9, which is only expressed in the epithelium, and not in the mesenchyme. FGF-10 helps to stimulate epithelial cell proliferation, in order make larger tooth germs. Mammalian teeth develop from ectoderm derived from the mesenchyme: oral ectoderm and neural crest. The epithelial components of the stem cells for continuously growing teeth form from tissue layers called the stellate reticulum and the suprabasal layer of the surface ectoderm.
Ectodermal dysplasia
Ectodermal dysplasia is a rare but severe condition where the tissue groups (specifically teeth, skin, hair, nails and sweat glands) derived from the ectoderm undergo abnormal development. Ectodermal dysplasia is a vague term, as there are over 170 subtypes of ectodermal dysplasia. It has been accepted that the disease is caused by a mutation or a combination of mutations in a number of genes. Research of the disease is ongoing, as only a fraction of the mutations involved with an ectodermal dysplasia subtype have been identified.
Hypohidrotic ectodermal dysplasia (HED) is the most common subtype of the disease. Clinical cases of patients with this condition displayed a range of symptoms. One of the common abnormalities of HED is hypohidrosis, or the inability to sweat, which can be attributed to dysfunctional sweat glands. This aspect can be especially dangerous in warm climates where the patient could potentially suffer from hyperthermia. Facial malformations are also related to HED such as disfigured or absent teeth, wrinkled skin around the eyes, misshaped nose along with scarce and thin hair. Skin problems, like eczema have also been observed in cases. It typically follows an X-linked recessive pattern of inheritance of the EDA genes. This disease typically affects males because they have only one X chromosome, meaning only one copy of the mutated gene is enough to cause abnormal development. For females to be affected, both X chromosomes would need to carry the gene mutation. If a female has a mutated version of the gene on one X chromosome, they are considered carrier of the disease.