
 | ||
In evolutionary biology, adaptive radiation is a process in which organisms diversify rapidly from an ancestral species into a multitude of new forms, particularly when a change in the environment makes new resources available, creates new challenges, or opens new environmental niches. Starting with a recent single ancestor, this process results in the speciation and phenotypic adaptation of an array of species exhibiting different morphological and physiological traits. An example of adaptive radiation would be the avian species of the Hawaiian honeycreepers. Via natural selection, these birds adapted rapidly and converged based on the different environments of the Hawaiian islands.
Contents
- Identification
- Innovation
- Darwins finches
- Cichlid fish
- Hawaiian honeycreepers
- Hawaiian silverswords
- Anolis lizards
- References
Much research has been done on adaptive radiation due to its dramatic effects on the diversity of a population. However, more research is needed, especially to fully understand the many factors affecting adaptive radiation. Both empirical and theoretical approaches are helpful, though each has its disadvantages. In order to procure the largest amount of data, empirical and theoretical approaches must be united.
Identification
Four features can be used to identify an adaptive radiation:
- A common ancestry of component species: specifically a recent ancestry. Note that this is not the same as a monophyly in which all descendants of a common ancestor are included.
- A phenotype-environment correlation: a significant association between environments and the morphological and physiological traits used to exploit those environments.
- Trait utility: the performance or fitness advantages of trait values in their corresponding environments.
- Rapid speciation: presence of one or more bursts in the emergence of new species around the time that ecological and phenotypic divergence is underway.
Innovation
The evolution of a novel feature may permit a clade to diversify by making new areas of morphospace accessible. A classic example is the evolution of a fourth cusp in the mammalian tooth. This trait permits a vast increase in the range of foodstuffs which can be fed on. Evolution of this character has thus increased the number of ecological niches available to mammals. The trait arose a number of times in different groups during the Cenozoic, and in each instance was immediately followed by an adaptive radiation. Birds find other ways to provide for each other, i.e. the evolution of flight opened new avenues for evolution to explore, initiating an adaptive radiation. Other examples include placental gestation (for eutherian mammals), or bipedal locomotion (in hominins).
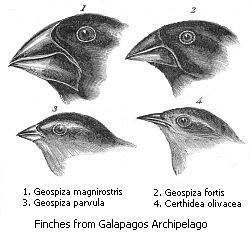
Darwin's finches
One famous example where adaptive radiation is seen is with Darwin's finches. It has been observed by many evolutionary biologists that fragmented landscapes are often a prime location for adaptive radiation to occur. The differences in geography throughout disjointed landscapes such as islands are believed to promote such diversification. Darwin's finches occupy the fragmented landscape of the Galápagos Islands and are diversified into many different species which differ in ecology, song, and morphology, specifically the size and shapes of their beaks. The first obvious explanation for these differences is allopatric speciation, speciation that occurs when populations of the same species become isolated geographically and evolve separately. Because the finches are divided amongst the islands, the birds have been evolving separately for several million years. However, this does not account for the fact that many of the species occur in sympatry, with seven or more species inhabiting the same island. This raises the question as to why these species split when living in the same environment with all the same resources. Petren, Grant, Grant, and Keller proposed that the speciation of the finches occurred in two parts: an initial, easily observable allopatric event followed by a less clear sympatric event. This sympatric event which occurred second was adaptive radiation. This occurred largely to promote specialization upon each island. One major morphological difference among species sharing one island is beak size and shape. Adaptive radiation led to the evolution of different beaks which could access different food and resources. Those with short beaks are better adapted to eating seeds on the ground, those with thin, sharp beaks eat insects, and those with long beaks use their beaks to probe for food inside cacti. With these specializations, seven or more species of finches are able to inhabit the same environments without competition or lack of resources killing several off. In other words, these morphological differences in beak size and shape brought about by adaptive radiation allow the island diversification to persist.
Cichlid fish
Another famous example is the cichlid fishes in lakes of the East African Rift. The lakes in this area are believed to support and sustain about 2,000 different species of these fish, each with different ecological and morphological characteristics such as body size. Like the Galápagos Islands, these lakes form a fragmented landscape that isolates the cichlid fish from one another, allowing them, and many of the organisms they live with, to evolve separately. The diversity of the lakes is in fact quite extraordinary because the adaptive radiations here are sometimes so young. One thing that has interested scientists about the cichlid fish case is the possibility of convergent evolution, or the evolution of analogous structures independently, driven by similar environmental selection pressures. However, quantitative studies on the convergent evolution of the cichlid fish are limited.
Hawaiian honeycreepers
Another example of an adaptive radiation would be an endemic species of the Hawaiian Islands. The Hawaiian honeycreepers are a large, highly diverse species which have been part of a vast adaptive radiation, that began as the Hawaiian Islands started to form. The honeycreeper species was shaped by island formation and natural selection. The mechanism by which this adaptive radiation occurred can be described as allopatric speciation via the peripheral isolate model. Each time a new island formed, a dispersal event would occur which would result in new community structures on each island. New selection pressures forced the adaptive radiation of the Hawaiian honeycreepers, as they needed to exploit new resources from the different environments of each island. It has been determined that many of the similar morphologies and behaviors of the Hawaiian Honeycreepers, located on distant islands, are due to convergence of analogous traits caused by similar environments.
Hawaiian silverswords
Though the most famously recognized cases of adaptive radiation have occurred in animals such as Darwin's finches or the cichlid fish, adaptive radiation certainly occurs in plant species as well. The most famous example of adaptive radiation in plants is quite possibly the Hawaiian silverswords. The Hawaiian silversword alliance consists of twenty-eight species of Hawaiian plants which range from trees to shrubs to vines. This is exceptional diversification as can be seen through the significant morphological differences between each species of the Hawaiian silverswords. With some species, it is virtually impossible to distinguish visually that they were ever part of one species to begin with. These radiations occurred millions of years ago, but through studies over the past few decades, it has been suggested that the rate of speciation and diversification was extremely high. These high rates, as well as the fragmented landscape of the Hawaiian Islands, are key characteristics which point directly to adaptive radiation.
Anolis lizards
Anolis lizards have been radiating widely in many different environments, including Central and South America, as well as the West Indies and experience great diversity of species just as the finches, cichlid fish, and silverswords. Studies have been done to determine whether radiations occur similarly for these lizards on the mainland as they do on the Caribbean islands or if differences can be observed in how they speciated. It has been observed that in fact, the radiations are very different, and ecological and morphological characteristics that these lizards developed as part of their speciation on the islands and on the mainland are unique. They have clearly evolved differently to the environments they inhabit. The environmental pressures on the Anolis lizards are not the same on the mainland as they are on the islands. There are significantly more predators preying on the Anolis lizards on the mainland. This is but one environmental difference. Other factors play a role in what sort of adaptive radiation will develop. Among the Caribbean islands, a larger perch diameter correlates with longer forelimbs, larger body mass, longer tails, and longer hind limbs. However, on the mainland, a larger perch diameter correlates with shorter tails. This shows that these lizards adapted differently to their environment depending on whether they were located on the mainland or the islands. These differing characteristics reconfirm that most of the adaptive radiation between the mainland and the islands occurred independently. On the islands specifically, species have adapted to certain "microhabitats" in which they require different morphological traits to survive. Irschick (1997) divides these microhabitats into six groups: "trunk–ground, trunk–crown, grass–bush, crown–giant, twig, and trunk." Different groups of lizards would acquire traits for one of these particular areas that made them more specialized for survival in this microhabitat and not so much in others. Adaptive radiation allows species to acquire the traits they need to survive in these microhabitats and reduce competition to allow the survival of a greater number of organisms as seen in many of the examples before.