
Symbol Sic1 UniProt P38634 | Entrez 850768 | |
 | ||
Alt. symbols YLR079W, SDB25, SIC1_YEAST, CDK inhibitor p40 | ||
Sic1, a protein, is a stoichiometric inhibitor of Cdk1-Clb (B-type cyclins) complexes in the budding yeast Saccharomyces cerevisiae. Because B-type cyclin-Cdk1 complexes are the drivers of S-phase initiation, Sic1 prevents premature S-phase entry. Multisite phosphorylation of Sic1 is thought to time Sic1 ubiquitination and destruction, and by extension, the timing of S-phase entry.
Contents
Role of Sic1 in the cell cycle control
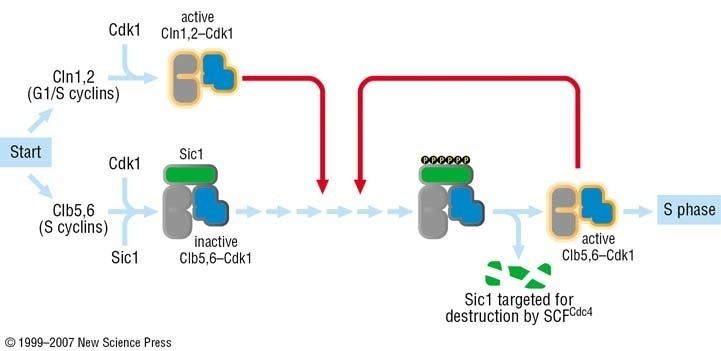
In the G1 phase of the cell cycle, Sic1 binds tightly to the Cdc28-Clb complex and inhibits it. Low Cdc28-Clb activity leads to the disassembly of the mitotic spindle, the assembly of the prereplicative complex and initiation of bud formation in yeast.
At the START point in the yeast cell cycle, the G1-cyclins Cln3, Cln1 and Cln 2 activate Cdc28. The activated complex will phosphorylate Sic1 at multiple sites which leads to its degradation by the SCF complex. When Sic1 is degraded, the Cdc28-Clb complex is no longer inhibited and the cell can enter the S/M-phase. Thus Sic1 inactivation is essential for transition into S phase (Fig.1).
Cdc28 in complex with B-type cyclin (Cdc28-Clb) phosphorylates Swi5, the transcription factor of Sic1. This promotes the export of Swi5 from the nucleus to the cytoplasm and avoids further transcription of the cdk inhibitor. Cdc28-Clb also phosphorylates any Sic1 molecules still available and triggers their ubiquitin-dependent degradation, exactly like Cdc28-Cln. High Cdc28-Clb levels also initiate DNA replication and duplication of the spindle pole bodies (SPBs). Then the metaphase spindle assembles and chromosome segregation can occur. The transcription of Sic1 starts during telophase, mediated by Swi5. Aca2 is another transcription factor of Sic1, but remains inactive until G1. At the end of mitosis, Sic1 is involved in the inactivation of Cdc28-Clb.
Ubiquitin-dependent degradation of Sic1
In order to be recognized by Cdc4 of the SCF complex, Sic1 has to be phosphorylated, often by Cyclin-Cdk complexes, at least at 6 of the 9 cdk sites (Fig. 2). Sic1 can also be phosphorylated by other kinases, such as Pho85-Pc11 , a kinase which becomes essential when Cln1 and Cln2 are absent. Sic1 has also a role in the response to osmostress. The stress-activated protein kinase (SAPK) Hog1 phosphorylates Sic1 at a single residue at the carboxyl terminus. This leads to downregulation of cyclin expression and Sic1 stabilization which arrests the cell cycle.
Multiple phosphorylations allow fine-tuning
Sic1 needs to be phosphorylated at multiple sites for ubiquitination-driven degradation (Fig. 2). The multiple phosphorylations are required for Sic1 to be recruited by Cdc4 to the SCF complex. The Cdc4 substrate recognition mechanism includes the interaction with consensus binding motifs on the surface of the folded and phosphorylated Sic1, the so-called Cdc4 phospho-degrons (CPD). It has been shown that the optimal consensus sequence for Cdc4 is a phosphorylated serine or threonine followed by a proline and a basic amino acid. However, none of the CPDs on the surface of the Sic1 show such a composition. Therefore, multiple phosphorylation of Sic1 is necessary to get high-affinity binding to Cdc4. Although this mechanism looks inefficient, it provides advantages for a cell because it is possible to measure the environmental Cln/cdc28 concentration. The number of phosphorylated sites corresponds to the concentration of Cln/cdc28 and Sic1 could be considered as a sensor for this protein. In contrast to the many sharp transitions of ultrasensitive kinase cascade feedback loops, this mechanism allows fine tuned regulation. Moreover, because multiple phosphorylations are required, the probability that Sic1 is degraded by random is small. Using multiple phosphorylation of Sic1, the cell evolved a strategy to highly regulate the onset of DNA replication that is absolutely vital to provide genetic stability.
Sic1 homologue in human and diseases
The protein p27Kip1 is a human homologue of Sic1, both having a conserved inhibitory domain, but p27Kip1 inhibits G1 cyclins and not cyclin B. There are several human diseases that are linked to p27Kip1 and other cyclin kinase inhibitors:
Thus, the human Cdk inhibitor p27Kip1 is a potential tumor suppressor protein. If its expression is reduced, the result might be unregulated progression from G1 to S-phase which deregulates cell division and simplifies the formation of tumors.