
Entrez 54205 | Ensembl ENSG00000172115 | |
 | ||
Aliases CYCS, CYC, HCS, THC4, cytochrome c, somatic, Cytochrome c, cyt c External IDs OMIM: 123970 MGI: 88578 HomoloGene: 133055 GeneCards: CYCS | ||
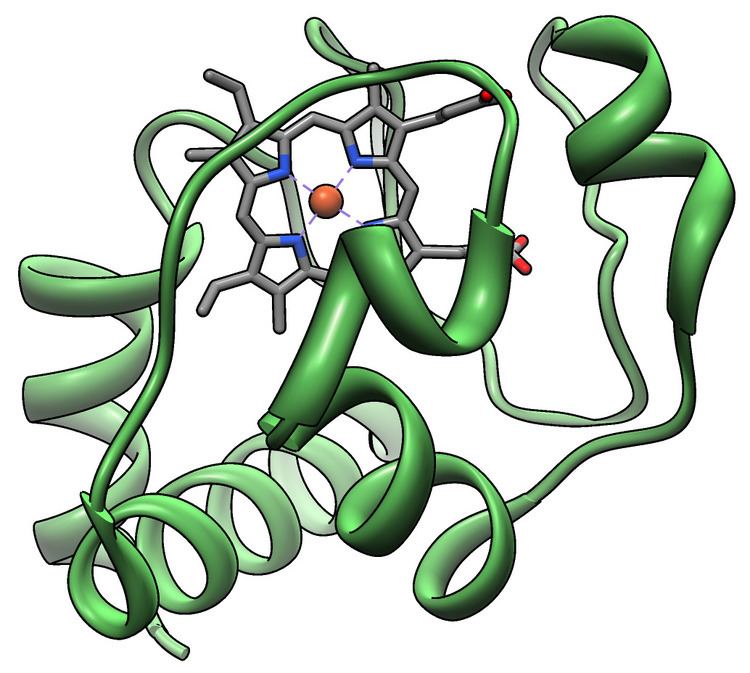
The cytochrome complex, or cyt c is a small hemeprotein found loosely associated with the inner membrane of the mitochondrion. It belongs to the cytochrome c family of proteins. Cytochrome c is highly water-soluble, unlike other cytochromes, and is an essential component of the electron transport chain, where it carries one electron. It is capable of undergoing oxidation and reduction, but does not bind oxygen. It transfers electrons between Complexes III (Coenzyme Q – Cyt C reductase) and IV (Cyt C oxidase). In humans, cytochrome c is encoded by the CYCS gene.
Contents
Species distribution
Cytochrome c is a highly conserved protein across the spectrum of species, found in plants, animals, and many unicellular organisms. This, along with its small size (molecular weight about 12,000 daltons), makes it useful in studies of cladistics. The cytochrome c molecule has been studied for the glimpse it gives into evolutionary biology.
Its primary structure consists of a chain of about 100 amino acids. Many higher-order organisms possess a chain of 104 amino acids. The sequences of cytochrome c in humans is identical to that of chimpanzees (our closest relatives), but differs more from that of horses.
Its amino acid sequence is highly conserved in eukaryotes, differing by only a few residues. In more than thirty species, 34 of the 104 amino acids are conserved; identical at that position. For example, human cytochrome oxidase reacts with wheat cytochrome c, in vitro; this is true for all pairs of species tested. In addition the redox potential of +0.25 volts is the same in all cytochrome c molecules studied.
Structure
All cytochrome c proteins contain a characteristic CXXCH (cysteine-any-any-cysteine-histidine) amino acid motif that binds heme. However, there are four classes of cytochrome c, each possessing a different fold.
Classes
In 1991 R. P. Ambler recognized four classes of cytochrome c:
Heme c
While most heme proteins are attached to the prosthetic group through iron ion ligation and tertiary interactions, the heme group of cytochrome c makes thioether bonds with two cysteine side chains of the protein. One of the main properties of heme c, which allows cytochrome c to have variety of functions, is its ability to have different reduction potentials in nature. This property determines the kinetics and thermodynamics of an electron transfer reaction.
Dipole moment
The dipole moment has an important role in orienting proteins to the proper directions and enhancing their abilities to bind to other molecules. The dipole moment of cytochrome c is a result from a cluster of negatively charged amino acid side chains at the "back" of the enzyme. Despite variations in the number of bound heme groups and variations in sequence, the dipole moment of vertebrate cytochromes c is remarkably conserved. For examples, vertebrate cytochromes c all have dipole moment of approximately 320 debye while cytochromes c of plants and insects have dipole moment of approximately 340 debye.
Function
Cytochrome c is a component of the electron transport chain in mitochondria. The heme group of cytochrome c accepts electrons from the bc1 complex and transfers electrons to the complex IV. Cytochrome c is also involved in initiation of apoptosis. Upon release of cytochrome c to the cytoplasm, the protein binds apoptotic protease activating factor-1 (Apaf-1).
Cytochrome c can also catalyze several redox reactions such as hydroxylation and aromatic oxidation, and shows peroxidase activity by oxidation of various electron donors such as 2,2-azino-bis(3-ethylbenzthiazoline-6-sulphonic acid) (ABTS), 2-keto-4-thiomethyl butyric acid and 4-aminoantipyrine.
A bacterial cytochrome c functions as a nitrite reductase.
Role in apoptosis
Cytochrome c is also an intermediate role in apoptosis, a controlled form of cell death used to kill cells in the process of development or in response to infection or DNA damage.
Cytochrome c binds to cardiolipin in the inner mitochondrial membrane, thus anchoring its presence and keeping it from releasing out of the mitochondria and initiating apoptosis. While the initial attraction between cardiolipin and cytochrome c is electrostatic due to the extreme positive charge on cytochrome c, the final interaction is hydrophobic, where a hydrophobic tail from cardiolipin inserts itself into the hydrophobic portion of cytochrome c.
During the early phase of apoptosis, mitochondrial ROS production is stimulated, and cardiolipin is oxidized by a peroxidase function of the cardiolipin–cytochrome c complex. The hemoprotein is then detached from the mitochondrial inner membrane and can be extruded into the soluble cytoplasm through pores in the outer membrane.
The sustained elevation in calcium levels precedes cyt c release from the mitochondria. The release of small amounts of cyt c leads to an interaction with the IP3 receptor (IP3R) on the endoplasmic reticulum (ER), causing ER calcium release. The overall increase in calcium triggers a massive release of cyt c, which then acts in the positive feedback loop to maintain ER calcium release through the IP3Rs. This explains how the ER calcium release can reach cytotoxic levels. This release of cytochrome c in turn activates caspase 9, a cysteine protease. Caspase 9 can then go on to activate caspase 3 and caspase 7, which are responsible for destroying the cell from within.
Inhibition of apoptosis
One of the ways cell apoptosis is activated is by release of cytochrome c from the mitochondria into cytosol. A study has shown that cells are able to protect themselves from apoptosis by blocking the release of cytochrome c using Bcl-xL. Another way that cells can control apoptosis is by phosphorylation of Tyr48 which would turn cytochrome c into an anti-apoptotic switch.
As an antioxidative enzyme
Cytochrome c is known to play a role in the electron transport chain and cell apoptosis. However, a recent study has shown that it can also act as an antioxidative enzyme in the mitochondria; and it does so by removing superoxide (O2–) and hydrogen peroxide (H2O2) from mitochondria. Therefore, not only is cytochrome c required in the mitochondria for cell respiration, but it is also needed in the mitochondria to limit the production of O2− and H2O2.
Extramitochondrial localization
Cytochrome c is widely believed to be localized solely in the mitochondrial intermembrane space under normal physiological conditions. The release of cytochrome-c from mitochondria to the cytosol, where it activates the caspase family of proteases is believed to be primary trigger leading to the onset of apoptosis. Measuring the amount of cytochrome c leaking from mitochondria to cytosol, and out of the cell to culture medium, is a sensitive method to monitor the degree of apoptosis. However, detailed immunoelectron microscopic studies with rat tissues sections employing cytochrome c-specific antibodies provide compelling evidence that cytochrome-c under normal cellular conditions is also present at extramitochondrial locations. In pancreatic acinar cells and the anterior pituitary, strong and specific presence of cytochrome-c was detected in zymogen granules and in growth hormone granules respectively. In the pancreas, cytochrome-c was also found in condensing vacuoles and in the acinar lumen. The extramitochondrial localization of cytochrome c was shown to be specific as it was completely abolished upon adsorption of the primary antibody with the purified cytochrome c. The presence of cytochrome-c outside of mitochondria at specific location under normal physiological conditions raises important questions concerning its cellular function and translocation mechanism. Besides cytochrome c, extramitochondrial localization has also been observed for large numbers of other proteins including those encoded by mitochondrial DNA. This raises the possibility about existence of yet-unidentified specific mechanisms for protein translocation from mitochondria to other cellular destinations.
Superoxide detection
Cytochrome c has been used to detect peroxide production in biological systems. As superoxide is produced, the number of oxidized cytochrome c3+ increases, and reduced cytochrome c2+ decreases. However, superoxide is often produced with nitric oxide. In the presence of nitric oxide, the reduction of cytochrome c3+ is inhibited. This leads to the oxidization of cytochrome c2+ to cytochrome c3+ by peroxynitrous acid, an intermediate made through the reaction of nitric oxide and superoxide. Presence of peroxynitrite, H2O2, or nitrogen dioxide NO2 in the mitochondria can be lethal since they nitrate tyrosine residues of cytochrome c which leads to disruption of cytochrome c’s function as an electron carrier in the electron transfer chain.
Low-level laser therapy
Cytochrome c is suspected to be the functional complex in so called LLLT: Low-level laser therapy. In LLLT, red light and some near infra-red wavelengths penetrate tissue in order to increase cellular regeneration. Light of this wavelength appears capable of increasing activity of cytochrome c, thus increasing metabolic activity and freeing up more energy for the cells to repair the tissue.