
 | ||
Cholera toxin (also known as choleragen and sometimes abbreviated to CTX, Ctx or CT) is protein complex secreted by the bacterium Vibrio cholerae. CTX is responsible for the massive, watery diarrhea characteristic of cholera infection.
Contents
History
Cholera toxin was discovered in 1959 by Late Prof.S.N.De (Ref.1) at Kolkata (India)
Structure
The cholera toxin is an oligomeric complex made up of six protein subunits: a single copy of the A subunit (part A, enzymatic), and five copies of the B subunit (part B, receptor binding), denoted as AB5. Subunit B binds while subunit A activates the G protein which activates adenylate cyclase. The three-dimensional structure of the toxin was determined using X-ray crystallography by Zhang et al. in 1995.
The five B subunits—each weighing 11 kDa, form a five-membered ring. The A subunit which is 28 kDa, has two important segments. The A1 portion of the chain (CTA1) is a globular enzyme payload that ADP-ribosylates G proteins, while the A2 chain (CTA2) forms an extended alpha helix which sits snugly in the central pore of the B subunit ring.
This structure is similar in shape, mechanism, and sequence to the heat-labile enterotoxin secreted by some strains of the Escherichia coli bacterium.
Pathogenesis
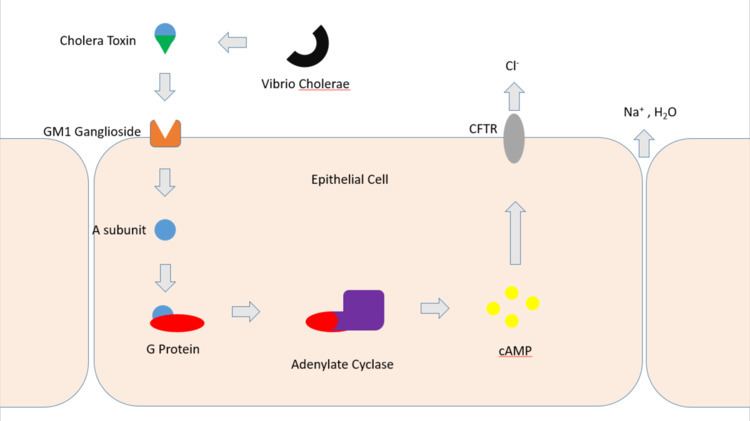
Cholera toxin acts by the following mechanism: First, the B subunit ring of the cholera toxin binds to GM1 gangliosides on the surface of target cells. The B subunit can also bind to cells lacking GM1. The toxin then most likely binds to other types of glycans attached to proteins instead of lipids. Once bound, the entire toxin complex is endocytosed by the cell and the cholera toxin A1 (CTA1) chain is released by the reduction of a disulfide bridge. The endosome is moved to the Golgi apparatus, where the A1 protein is recognized by the endoplasmic reticulum chaperon, protein disulfide isomerase. The A1 chain is then unfolded and delivered to the membrane, where Ero1 triggers the release of the A1 protein by oxidation of protein disulfide isomerase complex. As the A1 protein moves from the ER into the cytoplasm by the Sec61 channel, it refolds and avoids deactivation as a result of ubiquitination.
CTA1 is then free to bind with a human partner protein called ADP-ribosylation factor 6 (Arf6); binding to Arf6 drives a change in the shape of CTA1 which exposes its active site and enables its catalytic activity. The CTA1 fragment catalyses ADP-ribosylation of the Gs alpha subunit (Gαs) proteins using NAD. The ADP-ribosylation causes the Gαs subunit to lose its catalytic activity of inactivating GTP by hydrolyzing it to GDP + Pi, effectively increasing GTP concentration as less is being converted back to inactive GDP. Increased Gαs activation leads to increased adenylate cyclase activity, which increases the intracellular concentration of 3',5'-cyclic AMP (cAMP) to more than 100-fold over normal and over-activates cytosolic PKA. These active PKA then phosphorylate the cystic fibrosis transmembrane conductance regulator (CFTR) chloride channel proteins, which leads to ATP-mediated efflux of chloride ions and leads to secretion of H2O, Na+, K+, and HCO3− into the intestinal lumen. In addition, the entry of Na+ and consequently the entry of water into enterocytes are diminished. The combined effects result in rapid fluid loss from the intestine, up to 2 liters per hour, leading to severe dehydration and other factors associated with cholera, including a rice-water stool.
The pertussis toxin (also an AB5 protein) produced by Bordetella pertussis acts in a similar manner with the exception that it ADP-ribosylates the Gαi subunit, rendering it inactive and unable to inhibit adenylyl cyclase production of cAMP (leading to constitutive production).
Origin
The gene encoding the cholera toxin is introduced into V. cholerae by horizontal gene transfer. Virulent strains of V. cholerae carry a variant of temperate bacteriophage called the CTXf or CTXφ Bacteriophage.
Applications
Because the B subunit appears to be relatively non-toxic, researchers have found a number of applications for it in cell and molecular biology. It is routinely used as a neuronal tracer.
Treatment of cultured rodent neural stem cells with cholera toxin induces changes in the localization of the transcription factor Hes3 and increases their numbers.
GM1 gangliosides are found in lipid rafts on the cell surface. B subunit complexes labelled with fluorescent tags or subsequently targeted with antibodies can be used to identify rafts.