
 | ||
Trailer adaptation 2002
In biology, an adaptation, also called an adaptive trait, is a trait with a current functional role in the life of an organism that is maintained and evolved by means of natural selection. Adaptation refers to both the current state of being adapted and to the dynamic evolutionary process that leads to the adaptation. Adaptations enhance the fitness and survival of individuals. Organisms face a succession of environmental challenges as they grow and develop and are equipped with an adaptive plasticity as the phenotype of traits develop in response to the imposed conditions. The developmental norm of reaction for any given trait is essential to the correction of adaptation as it affords a kind of biological insurance or resilience to varying environments.
Contents
- Trailer adaptation 2002
- Adaptations in animals kindergarten preschoolers kids
- General principles
- Definitions
- Adaptedness and fitness
- Brief history
- Types of adaptations
- Changes in habitat
- Habitat tracking
- Genetic change
- Intimate relationships co adaptations
- Mimicry
- The basic machinery internal adaptations
- Compromise and conflict between adaptations
- Shifts in function
- Pre adaptations
- Co option of existing traits exaptation
- Non adaptive traits
- Vestigial organs
- Fitness landscapes
- Extinction
- Coextinction
- Flexibility acclimatization learning
- Flexibility
- Acclimatization
- Learning
- Diversity of genome DNAs
- Function and teleonomy
- Function
- Teleonomy
- References

Adaptations in animals kindergarten preschoolers kids
General principles
The significance of an adaptation can only be understood in relation to the total biology of the species.
Adaptation is, first of all, a process, to rather be with the animal and rather than a part of a body. An internal parasite (such as a liver fluke) can illustrate the distinction: such a parasite may have a very simple bodily structure, but nevertheless the organism is highly adapted to its specific environment. From this we see that adaptation is not just a matter of visible traits: in such parasites critical adaptations take place in the life cycle, which is often quite complex. However, as a practical term, "adaptation" often refers to a product: those features of a species which result from the process. Many aspects of an animal or plant can be correctly called adaptations, though there are always some features whose function remains in doubt. By using the term adaptation for the evolutionary process, and adaptive trait for the bodily part or function (the product), one may distinguish the two different senses of the word.
Adaptation is one of the two main processes that explain the diverse species found in biology, such as the different species of Darwin's finches. The other process is speciation (species-splitting or cladogenesis), caused by geographical isolation or some other mechanism. A favorite example used today to study the interplay of adaptation and speciation is the evolution of cichlid fish in African lakes, where the question of reproductive isolation is much more complex.

Adaptation is not always a simple matter where the ideal phenotype evolves for a given external environment. An organism must be viable at all stages of its development and at all stages of its evolution. This places constraints on the evolution of development, behavior and structure of organisms. The main constraint, over which there has been much debate, is the requirement that each genetic and phenotypic change during evolution should be relatively small, because developmental systems are so complex and interlinked. However, it is not clear what "relatively small" should mean, for example polyploidy in plants is a reasonably common large genetic change. The origin of eukaryotic symbiosis exemplifies a more dramatic example.
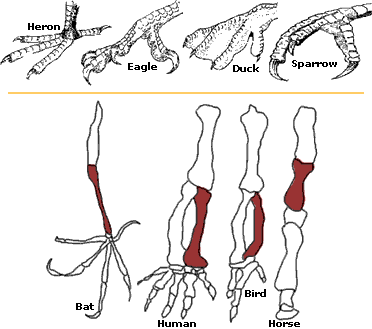
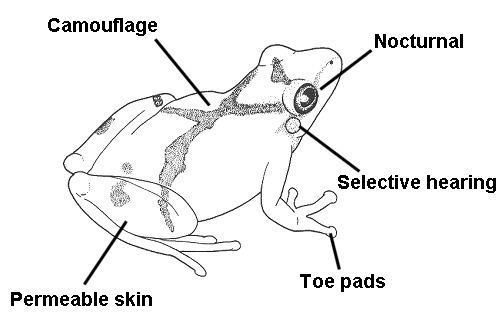
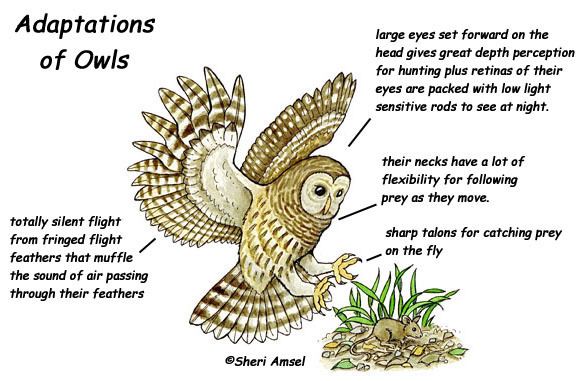
All adaptations help organisms survive in their ecological niches. The adaptive traits may be structural, behavioral or physiological. Structural adaptations are physical features of an organism (shape, body covering, armament; and also the internal organization). Behavioral adaptations are composed of inherited behavior chains and/or the ability to learn: behaviors may be inherited in detail (instincts), or a capacity for learning may be inherited (see neuropsychology). Examples: searching for food, mating, vocalizations. Physiological adaptations can permit the organism to perform special functions (for instance, making venom, secreting slime, phototropism); but may also involve more general functions such as growth and development, temperature regulation, ionic balance and other aspects of homeostasis. Adaptation, then, affects all aspects of the life of an organism.
Definitions
The following definitions are mainly due to Theodosius Dobzhansky.
1. Adaptation is the evolutionary process whereby an organism becomes better able to live in its habitat or habitats.2. Adaptedness is the state of being adapted: the degree to which an organism is able to live and reproduce in a given set of habitats.3. An adaptive trait is an aspect of the developmental pattern of the organism which enables or enhances the probability of that organism surviving and reproducing.Adaptedness and fitness
From the above definitions, it is clear that there is a relationship between adaptedness and fitness (a key population-genetics concept). Differences in fitness between genotypes predict the rate of evolution by natural selection. Natural selection changes the relative frequencies of alternative phenotypes, insofar as they are heritable. Although the two are connected, the one does not imply the other: a phenotype with high adaptedness may not have high fitness. Dobzhansky mentioned the example of the Californian redwood, which is highly adapted, but a relict species in danger of extinction. Elliott Sober commented that adaptation was a retrospective concept since it implied something about the history of a trait, whereas fitness predicts a trait's future.
1. Relative fitness. The average contribution to the next generation by a genotype or a class of genotypes, relative to the contributions of other genotypes in the population. This is also known as Darwinian fitness, selection coefficient, and other terms.2. Absolute fitness. The absolute contribution to the next generation by a genotype or a class of genotypes. Also known as the Malthusian parameter when applied to the population as a whole.3. Adaptedness. The extent to which a phenotype fits its local ecological niche. Researchers can sometimes test this through a reciprocal transplant.Brief history
Adaptation is a fact of life that has been accepted by many of the great thinkers who have tackled the world of living organisms. It is their explanations of how adaptation arises that separates these thinkers. A few of the most significant ideas:
Many other students of natural history, such as Buffon, accepted adaptation, and some also accepted evolution, without voicing their opinions as to the mechanism. This illustrates the real merit of Darwin and Alfred Russel Wallace, and secondary figures such as Henry Walter Bates, for putting forward a mechanism whose significance had only been glimpsed previously. A century later, experimental field studies and breeding experiments by people such as E. B. Ford and Dobzhansky produced evidence that natural selection was not only the 'engine' behind adaptation, but was a much stronger force than had previously been thought.
Types of adaptations
Adaptation is the heart and soul of evolution.
Changes in habitat
Before Charles Darwin, adaptation was seen as a fixed relationship between an organism and its habitat. It was not appreciated that as the climate changed, so did the habitat; and as the habitat changed, so did the biota. Also, habitats are subject to changes in their biota: for example, invasions of species from other areas. The relative numbers of species in a given habitat are always changing. Change is the rule, though much depends on the speed and degree of the change.
When the habitat changes, three main things may happen to a resident population: habitat tracking, genetic change or extinction. In fact, all three things may occur in sequence. Of these three effects only genetic change brings about adaptation.
Habitat tracking
When a habitat changes, the most common thing to happen is that the resident population moves to another locale which suits it; this is the typical response of flying insects or oceanic organisms, who have wide (though not unlimited) opportunity for movement. This common response is called habitat tracking. It is one explanation put forward for the periods of apparent stasis in the fossil record (the punctuated equilibrium theory).
Genetic change
Genetic change is what occurs in a population when natural selection acts on the genetic variability of the population; moreover, some mutations may create genetic variation that will lead to differing characteristics of offspring and hence abet adaptation. The first pathways of enzyme-based metabolism may have been parts of purine nucleotide metabolism, with previous metabolic pathways being part of the ancient RNA world. By this means, the population adapts genetically to its circumstances. Genetic changes may result in visible structures, or may adjust physiological activity in a way that suits the habitat.
It is now clear that habitats and biota do frequently change. Therefore, it follows that the process of adaptation is never finally complete. Over time, it may happen that the environment changes little, and the species comes to fit its surroundings better and better. On the other hand, it may happen that changes in the environment occur relatively rapidly, and then the species becomes less and less well adapted. Seen like this, adaptation is a genetic tracking process, which goes on all the time to some extent, but especially when the population cannot or does not move to another, less hostile area. Also, to a greater or lesser extent, the process affects every species in a particular ecosystem.
Leigh Van Valen thought that even in a stable environment, competing species had to constantly adapt to maintain their relative standing. This became known as the Red Queen hypothesis. One of the manifestations of the Red Queen dynamics can be seen in host-parasite interaction.
Intimate relationships: co-adaptations
In coevolution, where the existence of one species is tightly over bound up with the life of another species, new or 'improved' adaptations which occur in one species are often followed by the appearance and spread of corresponding features in the other species. There are many examples of this; the idea emphasises that the life and death of living things is intimately connected, not just with the physical environment, but with the life of other species. These relationships are intrinsically dynamic, and may continue on a trajectory for millions of years, as has the relationship between flowering plants and insects (pollination).
Pollinator constancy: these honey bees selectively visit flowers from only one species, as can be seen by the colour of the pollen in their baskets:
The gut contents, wing structures, and mouthpart morphologies of fossilized beetles and flies suggest that they acted as early pollinators. The association between beetles and angiosperms during the Early Cretaceous period led to parallel radiations of angiosperms and insects into the Late Cretaceous. The evolution of nectaries in Late Cretaceous flowers signals the beginning of the mutualism between hymenopterans and angiosperms.
Mimicry
Bates' work on Amazonian butterflies led him to develop the first scientific account of mimicry, especially the kind of mimicry which bears his name: Batesian mimicry. This is the mimicry by a palatable species of an unpalatable or noxious species. A common example seen in temperate gardens is the hoverfly, many of which—though bearing no sting—mimic the warning colouration of hymenoptera (wasps and bees). Such mimicry does not need to be perfect to improve the survival of the palatable species.
Bates, Wallace and Fritz Müller believed that Batesian and Müllerian mimicry provided evidence for the action of natural selection, a view which is now standard amongst biologists. All aspects of this situation can be, and have been, the subject of research. Field and experimental work on these ideas continues to this day; the topic connects strongly to speciation, genetics and development.
The basic machinery: internal adaptations
There are some important adaptations to do with the overall coordination of the systems in the body. Such adaptations may have significant consequences. Examples, in vertebrates, would be temperature regulation, or improvements in brain function, or an effective immune system. An example in plants would be the development of the reproductive system in flowering plants. Such adaptations may make the clade (monophyletic group) more viable in a wide range of habitats. The acquisition of such major adaptations has often served as the spark for adaptive radiation, and huge success over long periods of time for a whole group of animals or plants.
Compromise and conflict between adaptations
It is a profound truth that Nature does not know best; that genetical evolution... is a story of waste, makeshift, compromise and blunder.
All adaptations have a downside: horse legs are great for running on grass, but they can't scratch their backs; mammals' hair helps temperature, but offers a niche for ectoparasites; the only flying penguins do is under water. Adaptations serving different functions may be mutually destructive. Compromise and makeshift occur widely, not perfection. Selection pressures pull in different directions, and the adaptation that results is some kind of compromise.
Since the phenotype as a whole is the target of selection, it is impossible to improve simultaneously all aspects of the phenotype to the same degree.
Consider the antlers of the Irish elk, (often supposed to be far too large; in deer antler size has an allometric relationship to body size). Obviously, antlers serve positively for defence against predators, and to score victories in the annual rut. But they are costly in terms of resource. Their size during the last glacial period presumably depended on the relative gain and loss of reproductive capacity in the population of elks during that time. Another example: camouflage to avoid detection is destroyed when vivid colors are displayed at mating time. Here the risk to life is counterbalanced by the necessity for reproduction.
Stream-dwelling salamanders, such as Caucasian salamander or Gold-striped salamander have very slender, long bodies, perfectly adapted to life at the banks of fast small rivers and mountain brooks. Elongated body protects their larvae from being washed out by current. However, elongated body increases risk of desiccation and decreases dispersal ability of the salamanders; it also negatively affects their fecundity. As a result, fire salamander, less perfectly adapted to the mountain brook habitats, is in general more successful, have a higher fecundity and broader geographic range.
The peacock's ornamental train (grown anew in time for each mating season) is a famous adaptation. It must reduce his maneuverability and flight, and is hugely conspicuous; also, its growth costs food resources. Darwin's explanation of its advantage was in terms of sexual selection: "This depends on the advantage which certain individuals have over other individuals of the same sex and species, in exclusive relation to reproduction." The kind of sexual selection represented by the peacock is called 'mate choice,' with an implication that the process selects the more fit over the less fit, and so has survival value. The recognition of sexual selection was for a long time in abeyance, but has been rehabilitated. In practice, the Indian peafowl (Pavo cristatus) is a successful species, with a large natural range in India, so the overall outcome of their mating system is quite viable.
The conflict between the size of the human foetal brain at birth, (which cannot be larger than about 400 cm3, else it will not get through the mother's pelvis) and the size needed for an adult brain (about 1400 cm3), means the brain of a newborn child is quite immature. The most vital things in human life (locomotion, speech) just have to wait while the brain grows and matures. That is the result of the birth compromise. Much of the problem comes from our upright bipedal stance, without which our pelvis could be shaped more suitably for birth. Neanderthals had a similar problem.
As another example, the long neck of a giraffe is a burden and a blessing. The neck of a giraffe can be up to 2 m (6 ft 7 in) in length. This neck can be used for inter-species competition or for foraging on tall trees where shorter herbivores cannot reach. However, as previously stated, there is always a trade-off. This long neck is heavy and it adds to the body mass of a giraffe, so the giraffe needs an abundance of nutrition to provide for this costly adaptation.
Shifts in function
Adaptation and function are two aspects of one problem.
Pre-adaptations
This occurs when a species or population has characteristics which (by chance) are suited for conditions which have not yet arisen. For example, the polyploid cordgrass Spartina townsendii is better adapted than either of its parent species to their own habitat of saline marsh and mud-flats. White Leghorn chicken are markedly more resistant to vitamin B1 deficiency than other breeds. On a plentiful diet there is no difference, but on a restricted diet this preadaptation could be decisive.
Pre-adaptation may occur because a natural population carries a huge quantity of genetic variability. In diploid eukaryotes, this is a consequence of the system of sexual reproduction, where mutant alleles get partially shielded, for example, by the selective advantage of heterozygotes. Microorganisms, with their huge populations, also carry a great deal of genetic variability.
The first experimental evidence of the pre-adaptive nature of genetic variants in microorganisms was provided by Salvador Luria and Max Delbrück who developed Fluctuation Test, a method to show the random fluctuation of pre-existing genetic changes that conferred resistance to bacteriophage in the bacterium Escherichia coli.
Co-option of existing traits: exaptation
The classic example is the ear ossicles of mammals, which we know from paleontological and embryological studies originated in the upper and lower jaws and the hyoid bone of their synapsid ancestors, and further back still were part of the gill arches of early fish. We owe this esoteric knowledge to the comparative anatomists, who, a century ago, were at the cutting edge of evolutionary studies. The word exaptation was coined to cover these shifts in function, which are surprisingly common in evolutionary history. The origin of wings from feathers that were originally used for temperature regulation is a more recent discovery (see feathered dinosaurs).
Non-adaptive traits
Some traits do not appear to be adaptive, that is, they appear to have a neutral or even deleterious effect on fitness in the current environment. Because genes have pleiotropic effects, not all traits may be functional (i.e. spandrels). Alternatively, a trait may have been adaptive at some point in an organism's evolutionary history, but a change in habitats caused what used to be an adaptation to become unnecessary or even a hindrance (maladaptations). Such adaptations are termed vestigial.
Vestigial organs
Many organisms have vestigial organs, which are the remnants of fully functional structures in their ancestors. As a result of changes in lifestyle the organs became redundant, and are either not functional or reduced in functionality. With the loss of function goes the loss of positive selection, and the subsequent accumulation of deleterious mutations. Since any structure represents some kind of cost to the general economy of the body, an advantage may accrue from their elimination once they are not functional. Examples: wisdom teeth in humans; the loss of pigment and functional eyes in cave fauna; the loss of structure in endoparasites.
Fitness landscapes
Sewall Wright proposed that populations occupy adaptive peaks on a fitness landscape. In order to evolve to another, higher peak, a population would first have to pass through a valley of maladaptive intermediate stages. A given population might be "trapped" on a peak that is not optimally adapted.
Extinction
If a population cannot move or change sufficiently to preserve its long-term viability, then obviously, it will become extinct, at least in that locale. The species may or may not survive in other locales. Species extinction occurs when the death rate over the entire species exceeds the birth rate for a long enough period for the species to disappear. It was an observation of Van Valen that groups of species tend to have a characteristic and fairly regular rate of extinction.
Coextinction
Just as we have co-adaptation, there is also coextinction. Coextinction refers to the loss of a species due to the extinction of another; for example, the extinction of parasitic insects following the loss of their hosts. Coextinction can also occur when a flowering plant loses its pollinator, or through the disruption of a food chain. Ecologist Lian Pin Koh and colleagues discuss coextinction, stating, "Species coextinction is a manifestation of the interconnectedness of organisms in complex ecosystems. . . . While coextinction may not be the most important cause of species extinctions, it is certainly an insidious one."
Flexibility, acclimatization, learning
Flexibility deals with the relative capacity of an organism to maintain themselves in different habitats: their degree of specialization. Acclimatization is a term used for automatic physiological adjustments during life; learning is the term used for improvement in behavioral performance during life. In biology these terms are preferred, not adaptation, for changes during life which improve the performance of individuals. These adjustments are not inherited genetically by the next generation.
Adaptation, on the other hand, occurs over many generations; it is a gradual process caused by natural selection which changes the genetic make-up of a population so the collective performs better in its niche.
Flexibility
Populations differ in their phenotypic plasticity, which is the ability of an organism with a given genotype to change its phenotype in response to changes in its habitat, or to move to a different habitat.
To a greater or lesser extent, all living things can adjust to circumstances. The degree of flexibility is inherited, and varies to some extent between individuals. A highly specialized animal or plant lives only in a well-defined habitat, eats a specific type of food, and cannot survive if its needs are not met. Many herbivores are like this; extreme examples are koalas which depend on eucalyptus, and giant pandas which require bamboo. A generalist, on the other hand, eats a range of food, and can survive in many different conditions. Examples are humans, rats, crabs and many carnivores. The tendency to behave in a specialized or exploratory manner is inherited—it is an adaptation.
Rather different is developmental flexibility: "An animal or plant is developmentally flexible if when it is raised in or transferred to new conditions, it changes in structure so that it is better fitted to survive in the new environment," writes evolutionary biologist John Maynard Smith. Once again, there are huge differences between species, and the capacities to be flexible are inherited.
Acclimatization
If humans move to a higher altitude, respiration and physical exertion become a problem, but after spending time in high altitude conditions they acclimatize to the pressure by increasing production of Red blood cells. The ability to acclimatize is an adaptation, but not the acclimatization itself. Fecundity goes down, but deaths from some tropical diseases also goes down.
Over a longer period of time, some people will reproduce better at these high altitudes than others. They will contribute more heavily to later generations. Gradually the whole population becomes adapted to the new conditions. This we know takes place, because the performance of long-term communities at higher altitude is significantly better than the performance of new arrivals, even when the new arrivals have had time to make physiological adjustments.
Some kinds of acclimatization happen so rapidly that they are better called reflexes. The rapid colour changes in some flatfish, cephalopods, chameleons are examples.
Learning
Social learning is supreme for humans, and is possible for quite a few mammals and birds: of course, that does not involve genetic transmission except to the extent that the capacities are inherited. Similarly, the capacity to learn is an inherited adaptation, but not what is learnt; the capacity for human speech is inherited, but not the details of language.
Diversity of genome DNAs
A large diversity of genome DNAs in a species is the basis for species’ adaptation and for species’ differentiation. A great number of individuals are needed for carrying the great number of different genome DNAs. According to the Misrepair-accumulation aging theory, Misrepair mechanism is important in maintaining the sufficient number of individuals in a species. Misrepair is a way of repair for increasing the surviving chance of an organism when it has severe injuries. Without Misrepairs, no individual could survive to reproduction age. Thus Misrepair mechanism is an essential mechanism for the survival of a species and for maintaining the number of individuals. Although individuals die from aging, genome DNAs are being recopied and transmitted by individuals generation by generation. In addition, the DNA Misrepairs in germ cells contribute also to the diversity of genome DNAs.
Function and teleonomy
Adaptation raises some issues concerning how biologists use key terms such as function.
Function
To say something has a function is to say something about what it does for the organism, obviously. It also says something about its history: how it has come about. A heart pumps blood: that is its function. It also emits sound, which is just an ancillary side-effect. That is not its function. The heart has a history (which may be well or poorly understood), and that history is about how natural selection formed and maintained the heart as a pump. Every aspect of an organism that has a function has a history. Now, an adaptation must have a functional history: therefore we expect it must have undergone selection caused by relative survival in its habitat. It would be quite wrong to use the word adaptation about a trait which arose as a by-product.
It is widely regarded as unprofessional for a biologist to say something like "A wing is for flying," although that is their normal function. A biologist would be conscious that sometime in the remote past feathers on a small dinosaur had the function of retaining heat, and that later many wings were not used for flying (e.g. penguins, ostriches). So, the biologist would rather say that the wings on a bird or an insect usually had the function of aiding flight. That would carry the connotation of being an adaptation with a history of evolution by natural selection.
Teleonomy
Teleonomy is a term invented to describe the study of goal-directed functions which are not guided by the conscious forethought of man or any supernatural entity. It is contrasted with Aristotle's teleology, which has connotations of intention, purpose and foresight. Evolution is teleonomic; adaptation hoards hindsight rather than foresight. The following is a definition for its use in biology:
Teleonomy: The hypothesis that adaptations arise without the existence of a prior purpose, but by chance may change the fitness of an organism.The term may have been suggested by Colin Pittendrigh in 1958; it grew out of cybernetics and self-organising systems. Ernst Mayr, George C. Williams and Jacques Monod picked up the term and used it in evolutionary biology.
Philosophers of science have discussed the concept. Ernest Nagel analysed goal-directedness in biology; and David Hull commented on the use of teleology in biology:
...Haldane can be found remarking, 'Teleology is like a mistress to a biologist: he cannot live without her but he's unwilling to be seen with her in public.' Today the mistress has become a lawfully wedded wife. Biologists no longer feel obligated to apologize for their use of teleological language; they flaunt it. The only concession which they make to its disreputable past is to rename it 'teleonomy'.