
 | ||
Active transport is the movement of molecules across a cell membrane from a region of their lower concentration to a region of their higher concentration—in the direction against some gradient or other obstructing factor (often a concentration gradient).
Contents
- History
- Details
- Primary active transport
- Model of active transport
- Types of ATP powered primary active transporters
- Secondary active transport
- Antiport
- Symport
- Examples
- Endocytosis and Exocytosis
- References
Unlike passive transport, which uses the kinetic energy and natural entropy of molecules moving down a gradient, active transport uses cellular energy to move them against a gradient, polar repulsion, or other resistance. Active transport is usually associated with accumulating high concentrations of molecules that the cell needs, such as ions, glucose and amino acids. If the process uses chemical energy, such as from adenosine triphosphate (ATP), it is termed primary active transport. Secondary active transport involves the use of an electrochemical gradient. Examples of active transport include the uptake of glucose in the intestines in humans and the uptake of mineral ions into root hair cells of plants.
History
In 1848, the German physiologist Emil Heinrich du Bois-Reymond suggested the possibility of active transport of substances across membranes.
Rosenberg (1948) formulated the concept of active transport based on energetic considerations, but later it would be redefined.
Details
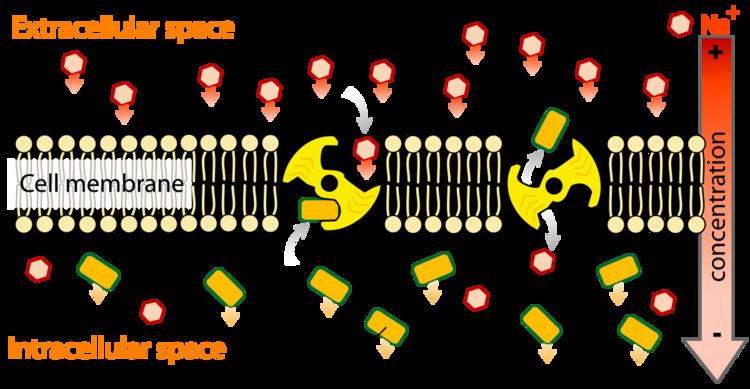
Specialized transmembrane proteins recognize the substance and allow it to move across the membrane when it otherwise would not, either because the phospholipid bilayer of the membrane is impermeable to the substance moved or because the substance is moved against the direction of its concentration gradient. There are two forms of active transport, primary active transport and secondary active transport. In primary active transport, the proteins involved are pumps that normally use the chemical energy in the form of ATP. Secondary active transport, however, makes use of potential energy, which are usually derived through exploitation of an electrochemical gradient. This involves pore-forming proteins that form channels across the cell membrane. The difference between passive transport and active transport is active transport requires energy and moves substances against their respective concentration gradient, whereas passive transport requires no energy and moves substances in the direction of their respective concentration gradient.
In an antiporter, one substrate is transported in one direction across the membrane while another is cotransported in the opposite direction. In a symporter, two substrates are transported in the same direction across the membrane. Antiport and symport processes are associated with secondary active transport, meaning that one of the two substances is transported in the direction of its concentration gradient, utilizing the energy derived from the transport of such substance (mostly Na+, K+ or H+ ions) down its concentration gradient.
If substrate molecules are moving from areas of lower concentration to areas of higher concentration (i.e., in the opposite direction as, or against the concentration gradient), specific transmembrane carrier proteins are required. These proteins have receptors that bind to specific molecules (e.g., glucose) and transport them across the cell membrane. Because energy is required in this process, it is known as 'active' transport. Examples of active transport include the transportation of sodium out of the cell and potassium into the cell by the sodium-potassium pump. Active transport often takes place in the internal lining of the small intestine.
Plants need to absorb mineral salts from the soil or other sources, but these salts exist in very dilute solution. Active transport enables these cells to take up salts from this dilute solution against the direction of the concentration gradient.
Primary active transport
Primary active transport, also called direct active transport, directly uses metabolic energy to transport molecules across a membrane.
Most of the enzymes that perform this type of transport are transmembrane ATPases. A primary ATPase universal to all animal life is the sodium-potassium pump, which helps to maintain the cell potential. The sodium-potassium pump maintains the membrane potential by moving three Na+ ions out of the cell for every two K+ ions moved into the cell. Other sources of energy for Primary active transport are redox energy and photon energy (light). An example of primary active transport using Redox energy is the mitochondrial electron transport chain that uses the reduction energy of NADH to move protons across the inner mitochondrial membrane against their concentration gradient. An example of primary active transport using light energy are the proteins involved in photosynthesis that use the energy of photons to create a proton gradient across the thylakoid membrane and also to create reduction power in the form of NADPH.
Model of active transport
ATP hydrolysis is used to transport hydrogen ions against the electrochemical gradient (from low to high hydrogen ion concentration). Phosphorylation of the carrier protein and the binding of a hydrogen ion induce a conformational (shape) change that drives the hydrogen ions to transport against the electrochemical gradient. Hydrolysis of the bound phosphate group and release of hydrogen ion then restores the carrier to its original conformation.
Types of ATP-powered primary active transporters
- P-type ATPase: sodium potassium pump, calcium pump, proton pump
- F-ATPase: mitochondrial ATP synthase, chloroplast ATP synthase
- V-ATPase: vacuolar ATPase
- ABC (ATP binding cassette) transporter: MDR, CFTR, etc.
Secondary active transport
In secondary active transport, also known as coupled transport or co-transport, energy is used to transport molecules across a membrane; however, in contrast to primary active transport, there is no direct coupling of ATP; instead it relies upon the electrochemical potential difference created by pumping ions in/out of the cell. Permitting one ion or molecule to move down an electrochemical gradient, but possibly against the concentration gradient where it is more concentrated to that where it is less concentrated increases entropy and can serve as a source of energy for metabolism (e.g. in ATP synthase).
In August 1960, in Prague, Robert K. Crane presented for the first time his discovery of the sodium-glucose cotransport as the mechanism for intestinal glucose absorption. Crane's discovery of cotransport was the first ever proposal of flux coupling in biology.
Cotransporters can be classified as symporters and antiporters depending on whether the substances move in the same or opposite directions.
Antiport
In an antiport two species of ion or other solutes are pumped in opposite directions across a membrane. One of these species is allowed to flow from high to low concentration which yields the entropic energy to drive the transport of the other solute from a low concentration region to a high one.
An example is the sodium-calcium exchanger or antiporter, which allows three sodium ions into the cell to transport one calcium out. Many cells also possess calcium ATPases, which can operate at lower intracellular concentrations of calcium and sets the normal or resting concentration of this important second messenger. But the ATPase exports calcium ions more slowly: only 30 per second versus 2000 per second by the exchanger. The exchanger comes into service when the calcium concentration rises steeply or "spikes" and enables rapid recovery. This shows that a single type of ion can be transported by several enzymes, which need not be active all the time (constitutively), but may exist to meet specific, intermittent needs.
Symport
Symport uses the downhill movement of one solute species from high to low concentration to move another molecule uphill from low concentration to high concentration (against its concentration gradient). Both molecules are transported in the same direction.
An example is the glucose symporter SGLT1, which co-transports one glucose (or galactose) molecule into the cell for every two sodium ions it imports into the cell. This symporter is located in the small intestines, heart, and brain. It is also located in the S3 segment of the proximal tubule in each nephron in the kidneys. Its mechanism is exploited in glucose rehydration therapy and defects in SGLT1 prevent effective reabsorption of glucose, causing familial renal glucosuria.
Examples
Endocytosis and Exocytosis
Endocytosis and exocytosis are both forms of bulk transport that move materials into and out of cells, respectively, via vesicles. In the case of Endocytosis, the cellular membrane folds around the desired materials outside the cell. The ingested particle becomes trapped within a pouch, known as a vesicle, inside the cytoplasm. Often enzymes from lysosomes are then used to digest the molecules absorbed by this process.
Biologists distinguish two main types of endocytosis: pinocytosis and phagocytosis.