
 | ||
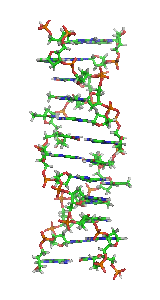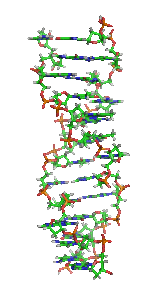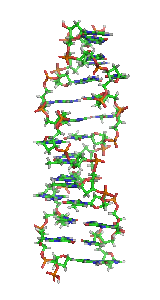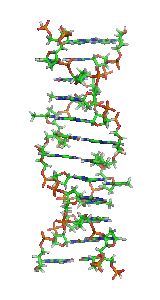
Z-DNA is one of the many possible double helical structures of DNA. It is a left-handed double helical structure in which the double helix winds to the left in a zig-zag pattern (instead of to the right, like the more common B-DNA form). Z-DNA is thought to be one of three biologically active double helical structures along with A- and B-DNA.
Contents
History
Left handed DNA was first discovered by Robert Wells and colleagues, during their studies of a repeating polymer of inosine-cytosine. They observed a "reverse" circular dichroism spectrum for such DNAs, and interpreted this (correctly) to mean that the strands wrapped around one another in a left handed fashion. The relationship between Z-DNA and the more familiar B-DNA was indicated by the work of Pohl and Jovin, who showed that the ultraviolet circular dichroism of poly(dG-dC) was nearly inverted in 4 M sodium chloride solution. The suspicion that this was the result of a conversion from B-DNA to Z-DNA was confirmed by examining the raman spectra of these solutions and the Z-DNA crystals. Subsequently, a crystal structure of "Z-DNA" was published which turned out to be the first single-crystal X-ray structure of a DNA fragment (a self-complementary DNA hexamer d(CG)3). It was resolved as a left-handed double helix with two anti-parallel chains that were held together by Watson-Crick base pairs (see: x-ray crystallography). It was solved by Andrew Wang, Alexander Rich, and co-workers in 1979 at MIT. The crystallisation of a B- to Z-DNA junction in 2005 provided a better understanding of the potential role Z-DNA plays in cells. Whenever a segment of Z-DNA forms, there must be B-Z junctions at its two ends, interfacing it to the B-form of DNA found in the rest of the genome.
In 2007, the RNA version of Z-DNA, Z-RNA, was described as a transformed version of an A-RNA double helix into a left-handed helix. The transition from A-RNA to Z-RNA, however, was already described in 1984.
Structure
Z-DNA is quite different from the right-handed forms. In fact, Z-DNA is often compared against B-DNA in order to illustrate the major differences. The Z-DNA helix is left-handed and has a structure that repeats every 2 base pairs. The major and minor grooves, unlike A- and B-DNA, show little difference in width. Formation of this structure is generally unfavourable, although certain conditions can promote it; such as alternating purine-pyrimidine sequence (especially poly(dGC)2), negative DNA supercoiling or high salt and some cations (all at physiological temperature, 37 °C, and pH 7.3-7.4). Z-DNA can form a junction with B-DNA (called a "B-to-Z junction box") in a structure which involves the extrusion of a base pair. The Z-DNA conformation has been difficult to study because it does not exist as a stable feature of the double helix. Instead, it is a transient structure that is occasionally induced by biological activity and then quickly disappears.
Predicting Z-DNA structure
It is possible to predict the likelihood of a DNA sequence forming a Z-DNA structure. An algorithm for predicting the propensity of DNA to flip from the B-form to the Z-form, ZHunt, was written by Dr. P. Shing Ho in 1984 (at MIT). This algorithm was later developed by Tracy Camp, P. Christoph Champ, Sandor Maurice, and Jeffrey M. Vargason for genome-wide mapping of Z-DNA (with P. Shing Ho as the principal investigator).
Biological significance
While no definitive biological significance of Z-DNA has been found, it is commonly believed to provide torsional strain relief (supercoiling) while DNA transcription occurs. The potential to form a Z-DNA structure also correlates with regions of active transcription. A comparison of regions with a high sequence-dependent, predicted propensity to form Z-DNA in human chromosome 22 with a selected set of known gene transcription sites suggests there is a correlation.
Toxic effect of ethidium bromide on trypanosomas is caused by shift of their kinetoplastid DNA to Z-form. The shift is caused by intercalation of EtBr and subsequent loosening of DNA structure that leads to unwinding of DNA, shift to Z-form and inhibition of DNA replication.
Z-DNA formed after transcription initiation
The first domain to bind Z-DNA with high affinity was discovered in ADAR1 using an approach developed by Alan Herbert. Crystallographic and NMR studies confirmed the biochemical findings that this domain bound Z-DNA in a non-sequence-specific manner. Related domains were identified in a number of other proteins through sequence homology. The identification of the Z-alpha domain provided a tool for other crystallographic studies that lead to the characterization of Z-RNA and the B-Z junction. Biological studies suggested that the Z-DNA binding domain of ADAR1 may localize this enzyme that modifies the sequence of the newly formed RNA to sites of active transcription.
In 2003, Alex Rich noticed that a poxvirus virulence factor, called E3L that has a Z-alpha related domain, mimicked a mammalian protein that binds Z-DNA. In 2005, Rich and his colleagues pinned down what E3L does for the poxvirus. When expressed in human cells, E3L increases by five- to 10-fold the production of several genes that block a cell's ability to self-destruct in response to infection.
Rich speculates that the Z-DNA is necessary for transcription and that E3L stabilizes the Z-DNA, thus prolonging expression of the anti-apoptotic genes. He suggests that a small molecule that interferes with the E3L binding to Z-DNA could thwart the activation of these genes and help protect people from pox infections.