
 | ||
The swim bladder, gas bladder, fish maw or air bladder is an internal gas-filled organ that contributes to the ability of many bony fish (but not cartilaginous fish) to control their buoyancy, and thus to stay at their current water depth without having to waste energy in swimming. Also, the dorsal position of the swim bladder means the center of mass is below the center of volume, allowing it to act as a stabilizing agent. Additionally, the swim bladder functions as a resonating chamber, to produce or receive sound.
Contents
- Structure and function
- Evolution
- Deep scattering layer
- Human uses
- Swim bladder disease
- Risk of injury
- Similar structures in other organisms
- References
The swim bladder is evolutionarily homologous to the lungs. Charles Darwin remarked upon this in On the Origin of Species. Darwin reasoned that the lung in air-breathing vertebrates had derived from a more primitive swim bladder, but scientists now believe that the swim bladder derived from a more primitive lung.
In the embryonic stages some species, such as redlip blenny, have lost the swim bladder again, mostly bottom dwellers like the weather fish. Other fish like the Opah and the Pomfret use their pectoral fins to swim and balance the weight of the head to keep a horizontal position. The normally bottom dwelling sea robin can use their pectoral fins to produce lift while swimming.
The gas/tissue interface at the swim bladder produces a strong reflection of sound, which is used in sonar equipment to find fish.
The cartilaginous fish (e.g. sharks and rays) do not have swim bladders. Some of them can control their depth only by swimming (using dynamic lift); others store fats or oils with density less than that of seawater to produce a neutral or near neutral buoyancy, which does not change with depth.
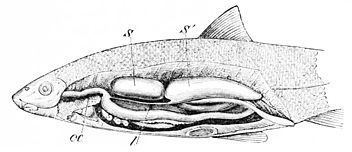
Structure and function
The swim bladder normally consists of two gas-filled sacs located in the dorsal portion of the fish, although in a few primitive species, there is only a single sac. It has flexible walls that contract or expand according to the ambient pressure. The walls of the bladder contain very few blood vessels and are lined with guanine crystals, which make them impermeable to gases. By adjusting the gas pressurising organ using the gas gland or oval window the fish can obtain neutral buoyancy and ascend and descend to a large range of depths. Due to the dorsal position it gives the fish lateral stability.
In physostomous swim bladders, a connection is retained between the swim bladder and the gut, the pneumatic duct, allowing the fish to fill up the swim bladder by "gulping" air. Excess gas can be removed in a similar manner.
In more derived varieties of fish (the physoclisti) the connection to the digestive tract is lost. In early life stages, these fish must rise to the surface to fill up their swim bladders; in later stages, the pneumatic duct disappears, and the gas gland has to introduce gas (usually oxygen) to the bladder to increase its volume and thus increase buoyancy. In order to introduce gas into the bladder, the gas gland excretes lactic acid and produces carbon dioxide. The resulting acidity causes the hemoglobin of the blood to lose its oxygen (Root effect) which then diffuses partly into the swim bladder. The blood flowing back to the body first enters a rete mirabile where virtually all the excess carbon dioxide and oxygen produced in the gas gland diffuses back to the arteries supplying the gas gland. Thus a very high gas pressure of oxygen can be obtained, which can even account for the presence of gas in the swim bladders of deep sea fish like the eel, requiring a pressure of hundreds of bars. Elsewhere, at a similar structure known as the oval window, the bladder is in contact with blood and the oxygen can diffuse back out again. Together with oxygen, other gases are salted out in the swim bladder which accounts for the high pressures of other gases as well.
The combination of gases in the bladder varies. In shallow water fish, the ratios closely approximate that of the atmosphere, while deep sea fish tend to have higher percentages of oxygen. For instance, the eel Synaphobranchus has been observed to have 75.1% oxygen, 20.5% nitrogen, 3.1% carbon dioxide, and 0.4% argon in its swim bladder.
Physoclist swim bladders have one important disadvantage: they prohibit fast rising, as the bladder would burst. Physostomes can "burp" out gas, though this complicates the process of re-submergence.
In some fish, mainly freshwater species (e.g. common carp, wels catfish), the swim bladder is connected to the labyrinth of the inner ear by the Weberian apparatus, a bony structure derived from the vertebrae, which provides a precise sense of water pressure (and thus depth), and improves hearing.
In red-bellied piranha, the swimbladder may play an important role in sound production as a resonator. The sounds created by piranhas are generated through rapid contractions of the sonic muscles and is associated with the swimbladder.
Evolution
Swim bladders are evolutionarily closely related (i.e., homologous) to lungs. It is believed that the first lungs, simple sacs connected to the gut that allowed the organism to gulp air under oxygen-poor conditions, evolved into the lungs of today's terrestrial vertebrates and some fish (e.g., lungfish, gar, and bichir) and into the swim bladders of the ray-finned fish. In embryonal development, both lung and swim bladder originate as an outpocketing from the gut; in the case of swim bladders, this connection to the gut continues to exist as the pneumatic duct in the more "primitive" ray-finned fish, and is lost in some of the more derived teleost orders. There are no animals which have both lungs and a swim bladder.
The cartilaginous fish (e.g., sharks and rays) split from the other fishes about 420 million years ago, and lack both lungs and swim bladders, suggesting that these structures evolved after that split. Correspondingly, these fish also have both heterocercal and pectoral fins which provide the necessary lift needed due to the lack of swim bladders. Teleost fish with swim bladders have neutral buoyancy, and have no need for this lift.
Deep scattering layer
Sonar operators, using the newly developed sonar technology during World War II, were puzzled by what appeared to be a false sea floor 300–500 metres deep at day, and less deep at night. This turned out to be due to millions of marine organisms, most particularly small mesopelagic fish, with swimbladders that reflected the sonar. These organisms migrate up into shallower water at dusk to feed on plankton. The layer is deeper when the moon is out, and can become shallower when clouds obscure the moon.
Most mesopelagic fish make daily vertical migrations, moving at night into the epipelagic zone, often following similar migrations of zooplankton, and returning to the depths for safety during the day. These vertical migrations often occur over large vertical distances, and are undertaken with the assistance of a swim bladder. The swim bladder is inflated when the fish wants to move up, and, given the high pressures in the mesoplegic zone, this requires significant energy. As the fish ascends, the pressure in the swimbladder must adjust to prevent it from bursting. When the fish wants to return to the depths, the swimbladder is deflated. Some mesopelagic fishes make daily migrations through the thermocline, where the temperature changes between 10 and 20 °C, thus displaying considerable tolerance for temperature change.
Sampling via deep trawling indicates that lanternfish account for as much as 65% of all deep sea fish biomass. Indeed, lanternfish are among the most widely distributed, populous, and diverse of all vertebrates, playing an important ecological role as prey for larger organisms. The estimated global biomass of lanternfish is 550–660 million metric tonnes, several times the entire world fisheries catch. Lanternfish also account for much of the biomass responsible for the deep scattering layer of the world's oceans. Sonar reflects off the millions of lanternfish swim bladders, giving the appearance of a false bottom.
Human uses
In some Asian cultures, the swim bladders of certain large fishes are considered a food delicacy. In China they are known as fish maw, 花膠/鱼鳔, and are served in soups or stews.
The vanity price of a vanishing kind of maw is behind the imminent extinction of the vaquita, the world's smallest dolphin breed. Only found in Mexico's Gulf of California, the once numerous vaquita now number less than 60 in total. Vaquita die in gillnets set to catch totoaba (the world's largest drum fish). Totoaba are being hunted to extinction for its maw, which can sell for as much $10,000 per kilogram.
Swim bladders are also used in the food industry as a source of collagen. They can be made into a strong, water-resistant glue, or used to make isinglass for the clarification of beer. In earlier times they were used to make condoms.
Swim bladder disease
Swim bladder disease is a common ailment in aquarium fish. A fish with swim bladder disorder can float nose down tail up, or can float to the top or sink to the bottom of the aquarium.
Risk of injury
Fish with a swim bladder are at greater risk of injury after exposure to loud impulsive sounds than those without.
Similar structures in other organisms
Siphonophores have a special swim bladder that allows the jellyfish-like colonies to float along the surface of the water while their tentacles trail below. This organ is unrelated to the one in fish.