
Scientific name Strepsiptera | Superorder Coleopterida Rank Order | |
 | ||
Similar Insect, Mecoptera, Embioptera, Zoraptera, Snakefly | ||
The peculiar strepsiptera life cycle
The Strepsiptera (translation: "twisted wing"', giving rise to the insects' common name, twisted-wing parasites) are an endopterygote order of insects with nine extant families making up about 600 species. The early-stage larvae and the short-lived adult males are not sessile, but most of their lives are spent as endoparasites in other insects, such as bees, wasps, leafhoppers, silverfish, and cockroaches.
Contents
- The peculiar strepsiptera life cycle
- Traumatic insemination and female counter adaptation in strepsiptera insecta
- Appearance and biology
- Classification
- Families
- Use in population control
- References
Traumatic insemination and female counter adaptation in strepsiptera insecta
Appearance and biology

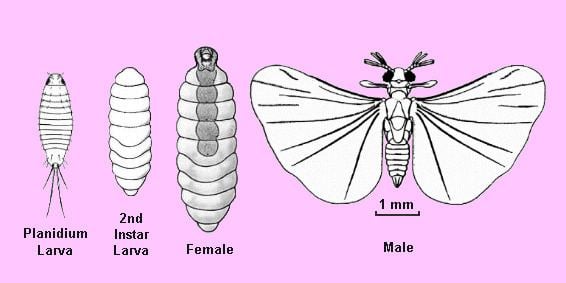
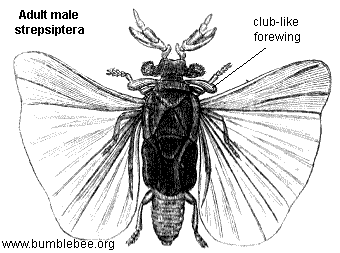
Males of the Strepsiptera have wings, legs, eyes, and antennae, and superficially look like flies, though their mouthparts cannot be used for feeding. Many of their mouthparts are modified into sensory structures. Adult males are very short-lived, usually surviving less than five hours, and do not feed. Females, in all families except the Mengenillidae, are not known to leave their hosts and are neotenic in form, lacking wings, legs, and eyes. Virgin females release a pheromone which the males use to locate them. In the Stylopidia, the female'S anterior region protrudes out of the host body and the male mates by rupturing the female's brood canal opening, which lies between the head and prothorax. Sperm passes through the opening in a process termed hypodermic insemination. The offspring consume their mother from the inside in a process known as hemocelous viviparity. Each female thus produces many thousands of triungulin larvae that emerge from the brood opening on the head, which protrudes outside the host body. These larvae have legs (which lack a trochanter, the leg segment that forms the articulation between the basal coxa and the femur), and actively search out new hosts. Their hosts include members of the orders Zygentoma, Orthoptera, Blattodea, Mantodea, Heteroptera, Hymenoptera, and Diptera. In the strepsipteran family Myrmecolacidae, the males parasitize ants, while the females parasitize Orthoptera.
Strepsiptera eggs hatch inside the female, and the planidium larvae can move around freely within the female's haemocoel, which is unique to these animals. The female has a brood canal that communicates with the outside world, and the larvae escape through this. The larvae are very active, as they only have a limited amount of time to find a host before they exhaust their food reserves. These first-instar larvae have stemmata (simple, single-lens eyes), and once they latch onto a host, they enter it by secreting enzymes that soften the cuticle, usually in the abdominal region of the host. Some species have been reported to enter the eggs of hosts. Larvae of Stichotrema dallatorreanurn Hofeneder from Papua New Guinea were found to enter their orthopteran host's tarsus (foot). Once inside the host, they undergo hypermetamorphosis and become a less mobile, legless larval form. They induce the host to produce a bag-like structure inside which they feed and grow. This structure, made from host tissue, protects them from the immune defences of the host. Larvae go through four more instars, and in each moult separation of the older cuticle occurs, but no discarding ("apolysis without ecdysis"), leading to multiple layers being formed around the larvae. Male larvae produce pupae after the last moult, but females directly become neotenous adults. The colour and shape of the host's abdomen may be changed and the host usually becomes sterile. The parasites then undergo pupation to become adults. Adult males emerge from the host bodies, while females stay inside. Females may occupy up to 90% of the abdominal volume of their hosts.

Adult male Strepsiptera have eyes unlike those of any other insect, resembling the schizochroal eyes found in the trilobite group known as the Phacopina. Instead of a compound eye consisting of hundreds to thousands of ommatidia, that each produce a pixel of the entire image - the strepsipteran eyes consist of only a few dozen "eyelets" that each produce a complete image. These eyelets are separated by cuticle and/or setae, giving the cluster eye as a whole a blackberry-like appearance.
Very rarely, multiple females may live within a single stylopized host; multiple males within a single host are somewhat more common. Adult males are rarely observed, however, although specimens may be lured using cages containing virgin females. Nocturnal specimens can also be collected at light traps.
Strepsiptera of the family Myrmecolacidae can cause their ant hosts to linger on the tips of grass leaves, increasing the chance of being found by the parasite's males (in case of females) and putting them in a good position for male emergence (in case of males).
Classification
The order, named by William Kirby in 1813, is named for the hind wings, which are held at a twisted angle when at rest (Greek στρεψι-, combining form of στρέϕειν, to twist; and πτερόν, wing). The fore wings are reduced to halteres (and initially were thought to be dried and twisted). Strepsiptera are an enigma to taxonomists. Originally, they were believed to be the sister group to the beetle families Meloidae and Ripiphoridae, which have similar parasitic development and forewing reduction; early molecular research suggested their inclusion as a sister group to the flies, in a clade called the halteria, which have one pair of the wings modified into halteres, and failed to support their relationship to the beetles. Further molecular studies, however, suggested they are outside the clade Mecopterida (containing the Diptera and Lepidoptera), but found no strong evidence for affinity with any other extant group. Study of their evolutionary position has been problematic due to difficulties in phylogenetic analysis arising from long branch attraction. The most basal strepsipteran is the fossil Protoxenos janzeni discovered in Baltic amber, while the most basal living strepsipteran is Bahiaxenos relictus, the sole member of the family Bahiaxenidae. The earliest known strepsipteran fossil is that of Cretostylops engeli, discovered in middle Cretaceous amber from Myanmar.
In 2012, a fresh molecular study revived the assertion that the Stepsiptera are the sister group of the Coleoptera (beetles).
Families
The Strepsiptera have two major groups: the Stylopidia and Mengenillidia. The Mengenillidia include three extinct families (Cretostylopidae, Protoxenidae, and Mengeidae) plus two extant families (Bahiaxenidae and Mengenillidae; the latter is not monophyletic, however.) They are considered more primitive, and the known females (Mengenillidae only) are free-living, with rudimentary legs and antennae. The females have a single genital opening. The males have strong mandibles, a distinct labrum, and more than five antennal segments.
The other group, the Stylopidia, includes seven families: the Corioxenidae, Halictophagidae, Callipharixenidae, Bohartillidae, Elenchidae, Myrmecolacidae, and Stylopidae. All Stylopidia have endoparasitic females having multiple genital openings.
The Stylopidae have four-segmented tarsi and four- to six-segmented antennae, with the third segment having a lateral process. The family Stylopidae may be paraphyletic. The Elenchidae have two-segmented tarsi and four-segmented antennae, with the third segment having a lateral process. The Halictophagidae have three-segmented tarsi and seven-segmented antennae, with lateral processes from the third and fourth segments. The Stylopidae mostly parasitize wasps and bees, the Elenchidae are known to parasitize Fulgoroidea, while the Halictophagidae are found on leafhoppers, treehoppers, and mole cricket hosts.
Strepsipteran insects in the genus Xenos parasitize Polistes carnifex, a species of social wasps. These obligate parasites infect the developing wasp larvae in the nest and are present within the abdomens of female wasps when they hatch out. Here they remain until they thrust through the cuticle and pupate (males) or release infective first-instar larvae onto flowers (females). These larvae are transported back to their nests by foraging wasps.
Use in population control
Many insects which have been considered as pests, such as leafhoppers and cockroaches, have species-specific Strepsipteran endoparasites. Inoculation of a pest population with the corresponding parasitoid has been found to aid in minimizing the impact of these pests. This method is useful in agriculture as a means of avoiding addition of chemicals to crops for pest control.