
 | ||
Lifespan Horse: 25 – 30 years, Cattle: 18 – 22 years Height Horse: 1.4 – 1.8 m, Human: 1.7 m Gestation period Horse: 11 – 12 months, Cattle: 283 days, Bonobo: 240 days, Common chimpanzee: 243 days Mass Horse: 380 – 1,000 kg, Cattle: 1,100 kg, Neanderthal: 78 kg Length Horse: 2.4 m, Bonobo: 70 – 83 cm, Common chimpanzee: 63 – 94 cm Speed Horse: 40 – 48 km/h, Human: 45 km/h Representative species Cattle, Human, Neanderthal, Bonobo, Upright man | ||
Viviparity literally means "giving live birth", from the Latin vivus ("living") and parere ("to beget"). The term is in use both in zoology and in botany as a term for certain classes of modes of reproduction, though it has different meanings in the two contexts.
The adjective viviparous is derived from "viviparity" and describes both the various modes of reproduction and the organisms that reproduce in any such mode.
Contents

In zoology viviparity refers to development of the embryo inside the body of the parent, eventually leading to live birth, as opposed to reproduction by laying eggs that complete their incubation outside the parental body. There are several forms of reproductive processes that answer to that description however, and they differ so radically, both in their nature and evolutionary origin, that no definitive and exclusive terminology is universally accepted, and the usage of the term is largely a matter of convenience and preference.

In botany vivipary is a common alternative form of the term, and botanical definitions also vary. One usage refers to reproduction via embryos, such as shoots or bulbils, as opposed to germinating externally from a dropped, dormant seed, as is usual in plants; however, vivipary also may refer to plants arising from buds or seedlings that have formed or germinated while still on the parent plant, and are dropped when ready to establish themselves in the substrate. In botany too, the usage of the term is largely a matter of convenience and preference.

In animals
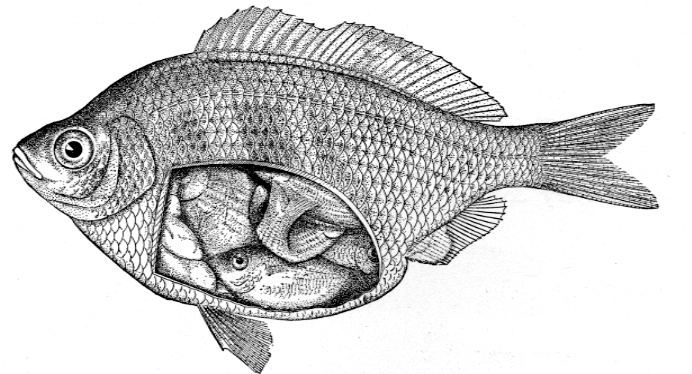
Five modes of reproduction have been differentiated in animals based on relations between zygote and parents. The five include two nonviviparous modes: ovuliparity, with external fertilisation, and oviparity, with internal fertilisation. In the latter, the female lays zygotes as eggs with a large yolk; this occurs in all birds, most reptiles, and some fishes. These modes are distinguished from viviparity, which covers all the modes that result in live birth:
At least some transport of nutrients from mother to embryo appears to be common to all viviparous species, but those with fully developed placentas such as found in the Theria, some skinks, and some fish can rely on the placenta for transfer of all necessary nutrients to the offspring and for removal of all the metabolic wastes as well once it has been fully established during the early phases of a pregnancy. In such species, there is direct, intimate contact between maternal and embryonic tissue, though there also is a placental barrier to control or prevent uncontrolled exchange and the transfer of pathogens.
In at least one species of skink in the large genus Trachylepis, placental transport accounts for nearly all of the provisioning of nutrients to the embryos before birth. In the uterus, the eggs are very small, about 1mm in diameter, with very little yolk and very thin shells. The shell membrane is vestigial and transient; its disintegration permits the absorption of nutrients from uterine secretions. The embryo then produces invasive chorionic tissues that grow between the cells of the uterine lining till they can absorb nutrients from maternal blood vessels. As it penetrates the lining, the embryonic tissue grows aggressively till it forms sheets of tissue beneath the uterine epithelium. They eventually strip it away and replace it, making direct contact with maternal capillaries. In several respects, the phenomenon is of considerable importance in theoretical zoology. The authors remark that such an endotheliochorial placenta is fundamentally different from that of any known viviparous reptile.
There is no relationship between sex-determining mechanisms and whether a species bears live young or lays eggs. Temperature-dependent sex determination, which cannot function in an aquatic environment, is seen only in terrestrial viviparous reptiles. Therefore, marine viviparous species, including sea snakes and, it now appears, the mosasaurs, ichthyosaurs, and plesiosaurs of the Cretaceous, use genotypic sex determination (sex chromosomes), much as birds and mammals do. Genotypic sex determination is also found in most reptiles, including many viviparous ones (such as Pseudemoia entrecasteauxii), whilst temperature dependent sex determination is found in some viviparous species, such as the montane water skink (Eulamprus tympanum).
In plants
Viviparous plants produce seeds or embryos that begin to develop before they detach from the parent. Plants such as some Iridaceae and Agavoideae grow cormlets in the axils of their inflorescences, These fall and in favourable circumstances they have effectively a whole season's start over fallen seeds. Similarly, some Crassulaceae, such as Bryophyllum, develop and drop plantlets from notches in their leaves, ready to grow. Such production of embryos from somatic issues is asexual vegetative reproduction that amounts to cloning.
Again, in some species of mangroves, for instance, the seed germinates and grows from its own resources while still attached to its parent. Seedlings of some species are dispersed by currents if they drop into the water, but others develop a heavy, straight taproot that commonly penetrates mud when the seedling drops, thereby effectively planting the seedling. This contrasts with the examples of vegetative reproduction mentioned above, in that the mangrove plantlets are true seedlings produced by sexual reproduction.
In some trees, like Jackfruit, some citrus, and avocado, the seeds can be found already germinated while the fruit goes overripe; strictly speaking this condition cannot be described as vivipary, but the moist and humid conditions provided by the fruit mimic a wet soil that encourages germination. However, the seeds also can germinate under moist soil.
Evolution
In general, viviparity and matrotrophy are believed to have evolved from an ancestral condition of oviparity and lecithotrophy (nutrients supplied through the yolk). One traditional hypothesis concerning the sequence of evolutionary steps leading to viviparity is a linear model. According to such a model, provided that fertilization was internal, the egg might have been retained for progressively longer periods in the reproductive tract of the mother. Through continued generations of egg retention, viviparous lecithotrophy may have gradually developed; in other words the entire development of the embryo, though still with nutrients provided by the yolk, occurred inside the mother’s reproductive tract, after which she would give birth to the young as they hatched. The next evolutionary development would be incipient matrotrophy, in which yolk supplies are gradually reduced and are supplemented with nutrients from the mother's reproductive tract.
In many ways, depending on the ecology and life strategy of the species, viviparity may be more strenuous and more physically and energetically taxing on the mother than oviparity. However, its numerous evolutionary origins imply that in some scenarios there must be worthwhile benefits to viviparous modes of reproduction; selective pressures have led to its convergent evolution more than 150 times among the vertebrates alone.
There is no one mode of reproduction that is universally superior in selective terms, but in many circumstances viviparity of various forms offers good protection from parasites and predators and permits flexibility in dealing with problems of reliability and economy in adverse circumstances. Variations on the theme in biology are enormous, ranging from trophic eggs to resorption of partly developed embryos in hard times or when they are too numerous for the mother to bring to term, but among the most profoundly advantageous features of viviparity are various forms of physiological support and protection of the embryo, such as thermoregulation and osmoregulation. Since the developing offspring remains within the mother’s body, she becomes, in essence, a walking incubator, protecting the developing young from excessive heat, cold, drought, or flood. This offers powerful options for dealing with excessive changes in climate or when migration events expose populations to unfavourable temperatures or humidities. In squamate reptiles in particular, there is a correlation between high altitudes or latitudes, colder climates and the frequency of viviparity. The idea that the tendency to favour egg-retention selectively under cooler conditions arises from the thermoregulatory benefits, and that it consequently promotes the evolution of viviparity as an adaptation, is known as "the cold climate hypothesis".